A ligand is an essential component of complex proteins. Concept of receptor and ligand
Topic: TRANSMEMBRANE TRANSFER
Intracellular receptors: https://www.youtube.com/watch?v=Nm9u4lNCPyM
Metabotropic membrane receptors associated with the second messenger system: https://www.youtube.com/watch?v=dQ4yVuLAbH0
Metabotropic membrane receptors associated with tyrosine kinase activity:
Types of transmembrane transport.
Types of transmembrane transport channels.
The concept of polar and non-polar substances.
Transmembrane selective permeability maintains cellular homeostasis, the optimal content of ions, water, enzymes and substrates in the cell. Ways to realize selective membrane permeability: passive transport, catalyzed transport (facilitated diffusion), active transport. The hydrophobic nature of the bilayer core determines the possibility (or impossibility) of direct penetration of substances various from a physicochemical point of view (primarily polar and nonpolar) through the membrane.
Non-polar substances(for example, cholesterol and its derivatives) freely penetrate biological membranes. For this reason, endocytosis and exocytosis of polar compounds (for example, peptide hormones) occur with the help of membrane vesicles, and the secretion of steroid hormones occurs without the participation of such vesicles. For the same reason, receptors for non-polar molecules (for example, steroid hormones) are located inside the cell.
Polar substances(eg proteins and ions) cannot penetrate biological membranes. This is why receptors for polar molecules (for example, peptide hormones) are built into plasma membrane, and signal transmission to other cellular compartments is carried out by second messengers. For the same reason, the transmembrane transfer of polar compounds is carried out by special systems built into biological membranes.
Selective permeability is provided by the cell membrane; The receptor function is realized by glycoproteins, the carbohydrate parts of which are located in the glycocalyx; shape retention and mobility are ensured by fibrillar and tubular proteins in the submembrane layer, etc.
Concept of receptor and ligand
Cell receptor- a molecule on the surface of the cell, nucleus, cellular organelles, or dissolved in the cytoplasm. The cellular receptor specifically reacts by changing its spatial configuration (shape) to the attachment of a molecule of a certain chemical substance to it - ligand, transmitting an external regulatory signal. This in turn transmits this signal into the cell or cell organelle. The place on the receptor where it attaches ligand is called a site. The same receptor can have several sites. Cellular receptors can be divided into two main classes - membrane receptors (located on the membrane separating the cell from the external environment) and intracellular receptors.
A substance that specifically binds to a receptor is called ligand (by messenger) this receptor. Thus, a ligand (synonym: messenger) is a chemical substance that can interact with a receptor. The outcome of this interaction may vary. If the ligand (messenger) leads to a change in shape receptor and its activation is called an agonist . If the ligand (messenger) changes the shape (conformation) of the receptor and block this receptor it is called an antagonist.
When it comes to the senses, ligands (messengers) are substances that act on the receptors of smell or taste.
There are also thermosensitive receptor proteins and receptor proteins that respond to changes in membrane potential.
Receptors for water-soluble ligands (messengers) - protein hormones, adrenaline, norepinephrine - are located on the surface of the membrane (membrane receptors), this is due to the fact that hydrophilic ligands cannot pass through the hydrophobic surface of the membrane. Fat-soluble ligands (messengers) easily pass through the phospholipid bilayer of the cell membrane and nucleus, and therefore the cell locates receptors (intracellular receptors) for them inside: on organelles, the nucleus. Examples of fat-soluble ligands can be steroid hormones of the adrenal glands and gonads.
In addition, ligands can be separated to exogenous(coming from outside) and endogenous(formed inside the body). As a rule, if some exogenous substance has receptors on cells, then the body also has endogenous ligands for this receptor. For example, the endogenous ligand of cannabinoid receptors to which cannabis alkaloids bind is the substance anandamide, produced by the body from arachidonic fatty acid. Or endorphin receptors (play an important role in the formation of pain and emotional state): can bind to endogenous ligands - endorphins, or can bind to drugs of the morphine group.
High specificity of binding of the protein to the ligand is ensured by the complementarity of the structure of the active center of the protein to the structure of the ligand
Complementarity refers to the spatial and chemical correspondence of interacting molecules. The ligand must have the ability to enter and spatially coincide with the conformation of the active site. This coincidence may not be complete, but due to the conformational lability of the protein, the active center is capable of small changes and is “adjusted” to the ligand. In addition, between the functional groups of the ligand and the amino acid radicals forming the active center, bonds must arise that hold the ligand in the active center. The bonds between the ligand and the active center of the protein can be either non-covalent (ionic, hydrogen, hydrophobic) or covalent.
1. Characteristics of the active center
The active center of a protein is a region relatively isolated from the environment surrounding the protein, formed by amino acid residues. In this region, each residue, due to its individual size and functional groups, forms the “relief” of the active center.
The combination of such amino acids into a single functional complex changes the reactivity of their radicals, just as the sound of a musical instrument in an ensemble changes. Therefore, the amino acid residues that make up the active center are often called an “ensemble” of amino acids.
The unique properties of the active center depend not only on the chemical properties of the amino acids that form it, but also on their precise relative orientation in space. Therefore, even minor violations of the general conformation of the protein as a result of point changes in its primary structure or conditions environment can lead to changes in the chemical and functional properties of the radicals that form the active center, disrupt the binding of the protein to the ligand and its function. During denaturation, the active center of proteins is destroyed and their biological activity is lost.
Often the active center is formed in such a way that the access of water to the functional groups of its radicals is limited, i.e. conditions are created for the binding of the ligand to amino acid radicals.
In some cases, the ligand attaches to only one of the atoms that has a certain reactivity, for example, the addition of O 2 to the iron of myoglobin or hemoglobin. However, the properties of a given atom to selectively interact with O2 are determined by the properties of the radicals surrounding the iron atom in the composition. Heme is also found in other proteins, such as cytochromes. However, the function of the iron atom in cytochromes is different; it serves as an intermediary for the transfer of electrons from one substance to another, while iron becomes either di- or trivalent.
The main property of proteins that underlies their functions is the selectivity of attachment of specific ligands to certain parts of the protein molecule.
2. Diversity of ligands
Ligands can be inorganic (often metal ions) and organic substances, low molecular weight and high molecular weight substances;
there are ligands that change their chemical structure when attached to the active site of a protein (changes in the substrate in the active site of the enzyme);
there are ligands that attach to the protein only at the time of functioning (for example, O 2 transported by hemoglobin), and ligands that are constantly associated with the protein and play an auxiliary role in the functioning of proteins (for example, iron, which is part of hemoglobin).
In cases where the amino acid residues that form the active center cannot ensure the functioning of a given protein, non-protein molecules can attach to certain areas of the active center. Thus, the active center of many enzymes contains a metal ion (cofactor) or an organic non-protein molecule (coenzyme). The non-protein part, firmly associated with the active center of the protein and necessary for its functioning, is called the “prostatic group”. Myoglobin, hemoglobin and cytochromes have a prosthetic group in the active center - heme, containing iron.
The connection of protomers in an oligomeric protein is an example of the interaction of high molecular weight ligands. Each protomer, connected to other protomers, serves as a ligand for them, just as they do for it.
Sometimes the attachment of a ligand changes the conformation of the protein, resulting in the formation of a binding site with other ligands. For example, the calmodulin protein, after binding to four Ca 2+ ions in specific areas, acquires the ability to interact with certain enzymes, changing their activity.
8. Quaternary structure of proteins. Features of the structure and functioning of oligomeric proteins using the example of hemoglobin. Cooperative changes in protomer conformation. Possibility of regulation of the biological function of oligomeric proteins by allosteric ligands.
By quaternary structure we mean the method of laying in space individual polypeptide chains that have the same (or different) primary, secondary or tertiary structure, and the formation of a structurally and functionally unified macromolecular formation. Many functional proteins consist of several polypeptide chains connected not by covalent bonds, but by non-covalent bonds (similar to those that ensure the stability of the tertiary structure). Each individual polypeptide chain, called a protomer, monomer or subunit, most often does not have biological activity. The protein acquires this ability through a certain method of spatial association of its constituent protomers, i.e. a new quality appears that is not characteristic of a monomeric protein. The resulting molecule is usually called an oligomer (or multimer). Oligomeric proteins are often built from an even number of protomers (from 2 to 4, less often from 6 to 8) with the same or different molecular weights - from several thousand to hundreds of thousands. In particular, the hemoglobin molecule consists of two identical α- and two β-polypeptide chains, i.e. is a tetramer.
Cooperative changes in protomer conformation.
A change in the conformation, and therefore the functional properties of all protomers of an oligomeric protein when a ligand is attached to only one of them, is called cooperative changes in the conformation of protomers.
Allosteric regulation . The enzyme modulates its activity with the help of an effector non-covalently bound to it. Binding occurs at a site spatially distant from the active (catalytic) center. This binding causes conformational changes in the protein molecule, leading to a change in the specific geometry of the catalytic center. Activity can increase - this is activation of the enzyme, or decrease - this is inhibition. The “message” about the addition of an allosteric activator is transmitted through conformational changes to the catalytic subunit, which becomes complementary to the substrate, and the enzyme “turns on”. When the activator is removed, the enzyme again goes into an inactive form and “turns off.” Allosteric regulation is the main way in which metabolic pathways are regulated.
Ppt%5C34928-slozhnye_belki_ch1_1.jpg" alt=">The active center of the protein and its interaction with the ligand. During the formation of the tertiary structure"> Активный центр белка и его взаимодействие с лигандом. В процессе формирования третичной структуры на поверхности функционально активного белка, обычно в углублении, образуется участок, сформированный радикалами аминокислот, далеко стоящими друг от друга в первичной структуре. Этот участок, имеющий уникальное строение для данного белка и способный специфично взаимодействовать с определенной молекулой или группой похожих молекул, называется центром связывания белка с лигандом или активным центром. Лигандами называются молекулы, взаимодействующие с белками.!}
Ppt%5C34928-slozhnye_belki_ch1_2.jpg" alt=">A ligand can be either a low molecular weight or a high molecular weight (macromolecule) substance, including"> Лигандом может быть как низкомолекулярное, так и высокомолекулярное (макромолекула) вещество, в том числе и другой белок. Лигандами являются субстраты ферментов, кофакторы, ингибиторы и активаторы ферментов, протомеры в олигомерном белке и т.д.!}
Ppt%5C34928-slozhnye_belki_ch1_3.jpg" alt=">High specificity of protein-ligand interaction is ensured by the complementarity of the structure of the active center to the structure of the ligand.">!}
Ppt%5C34928-slozhnye_belki_ch1_4.jpg" alt=">Complementarity is the spatial and chemical correspondence of interacting surfaces. The active center must not only"> Комплементарность - это пространственное и химическое соответствие взаимодействующих поверхностей. Активный центр должен не только пространственно соответствовать входящему в него лиганду, но и между функциональными группами радикалов, входящих в активный центр, и лигандом должны образоваться связи чаще всего нековалентные (ионные, водородные, а также гидрофобные взаимодействия), которые удерживают лиганд в активном центре.!}
Ppt%5C34928-slozhnye_belki_ch1_5.jpg" alt=">Complementary interaction of protein with ligand">!}
Ppt%5C34928-slozhnye_belki_ch1_6.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_7.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_8.jpg" alt="> CLASSIFICATION OF PROTEINS 1. Simple proteins consist only of amino acids. 2. Complex proteins (holoproteins)"> КЛАССИФИКАЦИЯ БЕЛКОВ 1. Простые белки состоят только из аминокислот. 2. Сложные белки (холопротеины) содержат белковую часть (апопротеин) и небелковую (простетическую) группу.!}
Ppt%5C34928-slozhnye_belki_ch1_9.jpg" alt=">Various organic (lipids, carbohydrates) and inorganic (metals) substances can act as a prosthetic group."> В качестве простетической группы могут выступать различные органические (липиды, углеводы) и неорганические (металлы) вещества. Связь между простетической группой и апопротеином может быть как ковалентная, так и нековалентная. Простетическую группу порой можно рассматривать в качестве лиганда. Наличие небелковой части обеспечивает выполнение белком его функции. При утрате простетической группы холопротеин теряет свою активность.!}
Ppt%5C34928-slozhnye_belki_ch1_10.jpg" alt=">Complex proteins - chromoproteins - nucleoproteins - lipoproteins - phosphoproteins - glycoproteins - metalloproteins">!}
Ppt%5C34928-slozhnye_belki_ch1_11.jpg" alt=">Metalloproteins include holoenzymes containing non-heme coordinated metal ions. Among metalloproteins there are proteins"> Металлопротеинам можно отнести холоферменты, содержащие негемовые координационно связанные ионы металлов. Среди металлопротеинов есть белки, выполняющие депонирующие и транспортные функции (например, железосодержащие ферритин и трансферрин) и ферменты (например, цинксодержащая карбоангидраза и различные супероксиддисмутазы, содержащие в качестве активных центров ионы меди, марганца, железа и других металлов). Но и хромопротеины, содержащие ионы металлов, также можно отнести к металлопротеинам.!}
Ppt%5C34928-slozhnye_belki_ch1_12.jpg" alt=">Metalloproteins are often enzymes. Metal ions in this case: - participate in the orientation of the substrate"> Металлопротеины часто являются ферментами. Ионы металлов в этом случае: - участвуют в ориентации субстрата в активном центре фермента, входят в состав активного центра фермента и участвуют в катализе, являясь, например, акцепторами электронов на определенной стадии ферментативной реакции. Часто ион металла в составе фермента называют кофактором.!}
Ppt%5C34928-slozhnye_belki_ch1_13.jpg" alt=">Enzymatic metalloproteins include proteins containing, for example: - copper - cytochrome oxidase, in the complex"> К ферментативным металлопротеинам относятся белки, содержащие например: - медь – цитохромоксидаза, в комплексе с другими ферментами дыхательной цепи митохондрий участвует в синтезе АТФ, - железо – ферритин, депонирующий железо в клетке, трансферрин, переносящий железо в крови, каталаза, обезвреживающая перекись водорода, - цинк – алкогольдегидрогеназа, обеспечивающая метаболизм этанола и других спиртов, лактатдегидрогеназа, участвующая в метаболизме молочной кислоты, - карбоангидраза, образующая угольную кислоту из CO2 и H2O, - щелочная фосфатаза, гидролизующая фосфорные эфиры различных соединений, - α2-макроглобулин, антипротеазный белок крови. - селен – тиреопероксидаза, участвующая в синтезе гормонов !} thyroid gland, antioxidant enzyme glutathione peroxidase, - calcium - α-amylase of saliva and pancreatic juice, hydrolyzing starch.
Ppt%5C34928-slozhnye_belki_ch1_14.jpg" alt=">Ferritin">!}
Ppt%5C34928-slozhnye_belki_ch1_15.jpg" alt=">Phosphoproteins are proteins that contain a phosphate group. It binds to the peptide chain"> Фосфопротеины – это белки, в которых присутствует фосфатная группа. Она связывается с пептидной цепью через остатки тирозина, серина и треонина, т.е. тех аминокислот, которые содержат ОН-группу. Способ присоединения фосфата к белку на примере серина и тирозина!}
Ppt%5C34928-slozhnye_belki_ch1_16.jpg" alt=">Phosphoric acid can perform: - A structural role, imparting charge, solubility and changing properties"> Фосфорная кислота может выполнять: - Структурную роль, придавая заряд, растворимость и изменяя свойства белка, например, в казеине молока, яичном альбумине. Наличие остатков фосфорной кислоты способствует связыванию кальция, что необходимо для формирования, например, костной ткани. - Функциональную роль. В клетке присутствует много белков, которые связаны с фосфатом не постоянно, а в зависимости от активности метаболизма. Белок может многократно переходить в фосфорилированную или в дефосфорилированную форму, что играет регулирующую роль в его работе.!}
Ppt%5C34928-slozhnye_belki_ch1_17.jpg" alt=">Phosphorylation is the process of transfer of a phosphoric acid residue from a phosphorylating donor agent to a substrate, usually"> Фосфорилирование - процесс переноса остатка фосфорной кислоты от фосфорилирующего агента-донора к субстрату, как правило, катализируемый ферментами (киназами) и ведущий к образованию эфиров фосфорной кислоты. Дефосфорилирование (утрату остатка фосфорной кислоты) катализируют фосфатазы. АТФ + R-OH → АДФ + R-OPO3H2 R-OPO3H2 + Н2О → R-OH + Н3РО4!}
Ppt%5C34928-slozhnye_belki_ch1_18.jpg" alt=">Examples: 1) enzymes glycogen synthase and glycogen phosphorylase 2) histones in the phosphorylated state bind less tightly"> Примеры: 1) ферменты гликогенсинтаза и гликогенфосфорилаза 2) гистоны в фосфорилированном состоянии менее прочно связываются с ДНК и активность генома возрастает. Изменение конформации белка в фосфорилированном и дефосфорилированном состоянии!}
Ppt%5C34928-slozhnye_belki_ch1_19.jpg" alt=">Lipoproteins contain non-covalently bound lipids as a prosthetic part. Lipids, in particular"> Липопротеины содержат в качестве простетической части нековалентно связанные липиды. Липиды, в частности жиры, холестерол и его эфиры не растворяются в водных фазах организма, поэтому транспорт их кровью и лимфой осуществляется в виде комплексов с белками и фосфолипидами, которые называются липопротеинами.!}
Ppt%5C34928-slozhnye_belki_ch1_20.jpg" alt=">All lipoproteins have a similar structure: the core consists of hydrophobic molecules: triacylglycerols, cholesterol esters, and"> Все липопротеины имеют сходное строение: ядро состоит из гидрофобных молекул: триацилглицеролов, эфиров холестерола, а на поверхности находится монослой фосфолипидов, полярные группы которых обращены к воде, а гидрофобные погружены в гидрофобное ядро липопротеина. Кроме фосфолипидов, на поверхности находятся белки – аполипопротеины (апобелками). Их выделяют несколько видов: А, В, С, D. В каждом типе липопротеинов преобладают соответствующие ему апобелки. Аполипопротеины выполняют различные функции. Интегральные аполипопротеины являются структурными компонентами. Периферические аполипопротеины в плазме крови могут передаваться от одного типа липопротеинов к другим, определяя их дальнейшие превращения.!}
Ppt%5C34928-slozhnye_belki_ch1_21.jpg" alt=">Scheme of the structure of a lipoprotein Structure of a lipoprotein">!}
Ppt%5C34928-slozhnye_belki_ch1_22.jpg" alt=">Structure of blood plasma lipoproteins">!}
Ppt%5C34928-slozhnye_belki_ch1_23.jpg" alt=">There are four main classes of lipoproteins: - high-density lipoproteins (HDL), - low-density lipoproteins (LDL),"> Выделяют четыре основных класса липопротеинов: -липопротеины высокой плотности (ЛПВП), -липопротеины низкой плотности (ЛПНП), -липопротеины очень низкой плотности (ЛПОНП), -хиломикроны (ХМ). Каждый из типов ЛП образуется в разных тканях и транспортирует определённые липиды. Концентрация и соотношение в крови тех или иных липопротеинов играют ведущую роль в возникновении такой распространенной сосудистой патологии как атеросклероз. ЛПВП являются антиатерогенными, ЛПНП и ЛПОНП – атерогенными.!}
Ppt%5C34928-slozhnye_belki_ch1_24.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_25.jpg" alt=">Glycoproteins or glycoconjugates are proteins containing a carbohydrate component covalently attached to a polypeptide backbone."> Гликопротеины или, гликоконъюгаты – это белки, содержащие углеводный компонент, ковалентно присоединенный к полипептидной основе. Содержание углеводов в них варьирует от 1% до 98% по массе. Два подкласса белков, содержащих углеводы: ■ протеогликаны ■ гликопротеины!}
Description="">
Ppt%5C34928-slozhnye_belki_ch1_27.jpg" alt=">Glycoproteins are characterized by a low content of carbohydrates, which are attached: - by an N-glycosidic bond to the NH2 group of some"> Для гликопротеинов характерно невысокое содержание углеводов, которые присоединены: - N-гликозидной связью к NН2-группе какого-нибудь аминокислотного остатка, например, аспарагина; - О-гликозидной связью к гидроксильной группе остатка серина, треонина,тирозина, гидроксилизина.!}
Ppt%5C34928-slozhnye_belki_ch1_28.jpg" alt=">Formation of O- and N-glycosidic bonds in glycoproteins. 1 - N-glycosidic bond between the amide group"> Образование О- и N-гликозидных связей в гликопротеинах. 1 - N-гликозидная связь между амидной группой аспарагина и ОН-группой моносахарида; 2 - О-гликозидная связь между ОН-группой серина и ОН-группой моносахарида.!}
Ppt%5C34928-slozhnye_belki_ch1_29.jpg" alt=">Method of adding carbohydrate to protein">!}
Ppt%5C34928-slozhnye_belki_ch1_30.jpg" alt=">The functions of glycoproteins are: 1. Structural - bacterial cell wall, bone matrix, for example, collagen, elastin."> Функцией гликопротеинов являются: 1. Структурная – клеточная стенка бактерий, костный матрикс, например, коллаген, эластин. 2. Защитная – например, антитела, интерферон, факторы свертывания крови (протромбин, фибриноген). 3. Рецепторная – присоединение эффектора приводит к изменению конформации белка-рецептора, что вызывает внутриклеточный ответ. 4. Гормональная – гонадотропный, адренокортикотропный и тиреотропный гормоны. 5. Ферментативная – холинэстераза, нуклеаза. 6. Транспортная – перенос веществ в крови и через мембраны, например, трансферрин, транскортин, альбумин, Na+,К+-АТФаза.!}
Ppt%5C34928-slozhnye_belki_ch1_31.jpg" alt=">Structure diagram of the receptor protein">!}
Ppt%5C34928-slozhnye_belki_ch1_32.jpg" alt=">Chromoproteins - collective name complex proteins with colored prosthetic groups of various chemical natures. "> Chromoproteins are the collective name for complex proteins with colored prosthetic groups of various chemical natures. hemoproteins (contain heme), retinalproteins (contain vitamin A), flavoproteins (contain vitamin B2), cobamide proteins (contain vitamin B12).
Ppt%5C34928-slozhnye_belki_ch1_33.jpg" alt=">Flavoproteins are enzymes of redox reactions. They contain vitamin B2 derivatives flavin mononucleotide (FMN) and flavin adenine dinucleotide"> Флавопротеины - это ферменты окислительно-восстановительных реакций. Содержат производные витамина В2 флавинмононуклеотид (ФМН) и флавинадениндинуклеотид (ФАД). Связываются данные простетические группы ковалентно и придают желтое окрашивание. Эти простетические группы являются производными изоаллоксазина.!}
Ppt%5C34928-slozhnye_belki_ch1_34.jpg" alt=">Isoalloxazine is a heterocyclic compound, a derivative of pteridine. The isoalloxazine molecule consists of three aromatic rings -"> Изоаллоксазин - гетероциклическое соединения, производное птеридина. Молекула изоаллоксазина состоит из трех ароматических колец - бензольного, пиримидинового, пиразинового.!}
Ppt%5C34928-slozhnye_belki_ch1_35.jpg" alt=">Hemoproteins are heme-containing chromoproteins. They include structurally similar iron or magnesium porphyrins as a non-protein component."> Гемопротеины - гем-содержащие хромопротеины. В качестве небелкового компонента включают структурно сходные железо- или магнийпорфирины. Белковый компонент может быть разнообразным как по составу, так и по структуре. Основу структуры простетической группы большинства гемосодержащих белков составляет порфириновое кольцо, являющееся в свою очередь производным тетрапиррольного соединения – порфирина. Порфирин!}
Ppt%5C34928-slozhnye_belki_ch1_36.jpg" alt=">The porphyrin ring is capable of forming coordination compounds with various metal ions. As a result of complexation,"> Порфириновое кольцо способно образовывать координационные соединения с различными ионами металлов. В результате комплексообразования формируются металлопорфирины: содержащие ионы железа – гемоглобины, миоглобин, цитохромы, пероксидаза, каталаза и др. (красное окрашивание), содержщие ионы магния – хлорофилл (зеленое окрашивание). Витамин В12 (кобалимин) содержит координированный ион кобальта Со2+ в порфириноподобном макроцикле – коррине, состоящем из четырех частично гидрированных пиррольных колец (розовое окрашивание).!}
Ppt%5C34928-slozhnye_belki_ch1_37.jpg" alt=">Chlorophyll b. Chlorophylls are involved in the processes of photosynthesis.">!}
Ppt%5C34928-slozhnye_belki_ch1_38.jpg" alt=">Cytochromes differ in the amino acid composition of the peptide chains, the number of chains and are divided into types a, b,"> Цитохромы различаются аминокислотным составом пептидных цепей, числом цепей и разделяются на типы а, b, с, d. Цитохромы находятся в составе дыхательной цепи и цепи микросомального окисления. Степень окисления железа в составе цитохромов меняется в отличие от гемоглобина и миоглобина Fe2+ ↔ Fe3+!}
Ppt%5C34928-slozhnye_belki_ch1_39.jpg" alt=">Myoglobin (MB) is a protein found in red muscles, the main function of which is to create reserves"> Миоглобин (Мв) - белок, находящийся в красных мышцах, основная функция которого - создание запасов О2, необходимых при интенсивной мышечной работе. Мв - сложный белок, содержащий белковую часть - апоМв и небелковую часть - гем. Первичная структура апоМв определяет его компактную глобулярную конформацию и структуру активного центра, к которому присоединяется небелковая часть миоглобина - гем. Кислород, поступающий из крови в мышцы, связывается с Fe2+ гема в составе миоглобина. Мв - мономерный белок, имеющий очень высокое сродство к О2, поэтому отдача кислорода миоглобином происходит только при интенсивной мышечной работе, когда парциальное давление O2 резко снижается. Формирование пространственных структур и функционирование миоглобина.!}
Ppt%5C34928-slozhnye_belki_ch1_40.jpg" alt=">Formation of the Mv conformation. In red muscles, synthesis of the primary"> Формирование конформации Мв. В красных мышцах на рибосомах в ходе трансляции идет синтез первичной структуры Мв, представленной специфической последовательностью 153 аминокислотных остатков. Вторичная структура Мв содержит восемь α-спиралей, называемых латинскими буквами от А до Н, между которыми имеются неспирализованные участки. Третичная структура Мв имеет вид компактной глобулы, в углублении которой между F и Е α-спиралями расположен активный центр.!}
Ppt%5C34928-slozhnye_belki_ch1_41.jpg" alt=">Structure of myoglobin">!}
Ppt%5C34928-slozhnye_belki_ch1_42.jpg" alt=">Features of the structure and functioning of the active center of Mv. The active center of Mv is formed predominantly by hydrophobic radicals"> Особенности строения и функционирования активного центра Мв. Активный центр Мв сформирован преимущественно гидрофобными радикалами аминокислот, далеко отстоящими друг от друга в первичной структуре (например, Три39 и Фен138). К активному центру присоединяется плохо растворимые в воде лиганды - гем и О2. Гем - специфический лиганд апоМв.!}
Ppt%5C34928-slozhnye_belki_ch1_43.jpg" alt=">The basis of heme is made up of four pyrrole rings connected by methyl bridges; in the center there is a Fe2+ atom,"> Основу гема составляют четыре пиррольных кольца, соединенных метенильными мостиками; в центре расположен атом Fe2+, соединенный с атомами азота пиррольных колец четырьмя координационными связями. В активном центре Мв кроме гидрофобных радикалов аминокислот имеются также остатки двух аминокислот с гидрофильными радикалами - Гис Е7 (Гис64) и Гис F8 (Гис93).!}
Ppt%5C34928-slozhnye_belki_ch1_44.jpg" alt=">His F8 forms a coordination bond with Fe2+ and firmly fixes heme in the active site."> Гис F8 образует координационную связь с Fe2+ и прочно фиксирует гем в активном центре. Гис Е7 необходим для правильной ориентации в активном центре другого лиганда - O2 при его взаимодействии с Fe+2 гема. Микроокружение гема создает условия для прочного, но обратимого связывания O2 с Fe+2 и препятствует попаданию в гидрофобный активный центр воды, что может привести к его окислению в Fе3+.!}
Ppt%5C34928-slozhnye_belki_ch1_45.jpg" alt=">Oligomeric structure of HB and regulation of HB affinity for O2 by ligands. Human hemoglobins -"> Олигомерное строение Нв и регуляция сродства Нв к О2 лигандами. Гемоглобины человека - семейство белков, так же как и миоглобин относящиеся к сложным белкам (гемопротеинам). Они имеют тетрамерное строение и содержат две α-цепи, но различаются по строению двух других полипептидных цепей (2α-, 2х-цепи). Строение второй полипептидной цепи определяет особенности функционирования этих форм Нв. Около 98% гемоглобина эритроцитов взрослого человека составляет гемоглобин А (2α-, 2β-цепи). В период внутриутробного развития функционируют два основных типа гемоглобинов: эмбриональный Нв (2α, 2ε), который обнаруживается на ранних этапах развития плода, и гемоглобин F (фетальный) - (2α, 2γ), который приходит на смену раннему гемоглобину плода на шестом месяце внутриутробного развития и только после рождения замещается на Нв А.!}
Ppt%5C34928-slozhnye_belki_ch1_46.jpg" alt=">Hb A is a protein related to myoglobin (Mb), found in the red blood cells of an adult. Its structure"> Нв А - белок, родственный миоглобину (Мв), содержится в эритроцитах взрослого человека. Строение его отдельных протомеров аналогично таковому у миоглобина. Вторичная и третичная структуры миоглобина и протомеров гемоглобина очень сходны, несмотря на то что в первичной структуре их полипептидных цепей идентичны только 24 аминокислотных остатка (вторичная структура протомеров гемоглобина, так же как миоглобин, содержит восемь α-спиралей, обозначаемых латинскими буквами от А до Н, а третичная структура имеет вид компактной глобулы). Но в отличие от миоглобина гемоглобин имеет олигомерное строение, состоит из четырех полипептидных цепей, соединенных нековалентными связями.!}
Ppt%5C34928-slozhnye_belki_ch1_47.jpg" alt=">Oligomeric structure of hemoglobin">!}
Ppt%5C34928-slozhnye_belki_ch1_48.jpg" alt=">Each Hb protomer is associated with a non-protein part - heme and neighboring protomers. Connection of protein"> Каждый протомер Нв связан с небелковой частью - гемом и соседними протомерами. Соединение белковой части Нв с гемом аналогично таковому у миоглобина: в активном центре белка гидрофобные части гема окружены гидрофобными радикалами аминокислот за исключением Гис F8 и Гис Е7, которые расположены по обе стороны от плоскости гема и играют аналогичную роль в функционировании белка и связывании его с кислородом. Кроме того, Гис Е7 выполняет важную дополнительную роль в функционировании Нв. Свободный гем имеет в 25 000 раз более высокое сродство к СО, чем к О2. СО в небольших количествах образуется в организме и, учитывая его высокое сродство к гему, он мог бы нарушать транспорт необходимого для жизни клеток О2. Однако в составе гемоглобина сродство гема к оксиду углерода превышает сродство к О2 всего в 200 раз благодаря наличию в активном центре Гис Е7. Остаток этой аминокислоты создает оптимальные условия для связывания гема с O2 и ослабляет взаимодействие гема с СО.!}
Ppt%5C34928-slozhnye_belki_ch1_49.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_50.jpg" alt=">The pyrrole rings of heme are located in the same plane, and the Fe2+ ion is in the non-oxygenated state Hb"> Пиррольные кольца гема расположены в одной плоскости, а ион Fe2+ в неоксигенированом состоянии Hb выступает над плоскостью на 0,6 А. При присоединении кислорода ион железа погружается в плоскость колец гема. В результате сдвигается и участок полипептидной цепи, нарушаются слабые связи в молекуле Hb и изменяется конформация всей глобулы. Таким образом, присоединение кислорода вызывает изменение пространственной структуры молекулы миоглобина или протомеров гемоглобина.!}
Ppt%5C34928-slozhnye_belki_ch1_51.jpg" alt=">Hemoglobin can exist both in free (deoxyhemoglobin) and in oxygenated form, adding up to"> Гемоглобин может существовать как в свободной (дезоксигемоглобин), так и в оксигенированной форме, присоединяя до 4 молекул кислорода. Взаимодействие с кислородом 1-го протомера вызывает изменение его конформации, а также кооперативные конформационные изменения остальных протомеров. Сродство к кислороду возрастает, и присоединение О2 к активному центру 2-го протомера происходит легче, вызывая дальнейшую конформационную перестройку всей молекулы. В результате еще сильнее изменяется структура оставшихся протомеров и их активных центров, взаимодействие с О2 еще больше облегчается. В итоге 4-я молекула кислорода присоединяется к Hb примерно в 300 раз легче, чем 1-я. Так происходит в легких при высоком парциальном давлении кислорода.!}
Ppt%5C34928-slozhnye_belki_ch1_52.jpg" alt=">Cooperative changes in the conformation of the hemoglobin molecule upon interaction with oxygen">!}
Ppt%5C34928-slozhnye_belki_ch1_53.jpg" alt=">In tissues where the oxygen content is lower, on the contrary, the cleavage of each O2 molecule facilitates the release of subsequent ones."> В тканях, где содержание кислорода ниже, наоборот, отщепление каждой молекулы О2 облегчает освобождение последующих. Таким образом, взаимодействие олигомерного белка гемоглобина с лигандом (О2) в одном центре связывания приводит к изменению конформации всей молекулы и других, пространственно удаленных центров, расположенных на других субъединицах (принцип «домино»). Подобные взаимосвязанные изменения структуры белка называют кооперативными конформационными изменениями. Они характерны для всех олигомерных белков и используются для регуляции их активности.!}
Ppt%5C34928-slozhnye_belki_ch1_54.jpg" alt=">The interaction of both proteins (Mb and Hb) with oxygen depends on its partial pressure in"> Взаимодействие обоих белков (Mb и Hb) с кислородом зависит от его парциального давления в тканях. Эта зависимость имеет разный характер, что связано с их особенностями структуры и функционирования. Гемоглобин имеет S-образную кривую насыщения, которая показывает, что субъединицы белка работают кооперативно, и чем больше кислорода они отдают, тем легче идет освобождение остальных молекул О2. Этот процесс зависит от изменения парциального давления кислорода в тканях. График насыщения миоглобина кислородом имеет характер простой гиперболы, т.е. насыщение Mb кислородом происходит быстро и отражает его функцию - обратимое связывание с кислородом, высвобождаемым гемоглобином, и освобождение в случае интенсивной физической нагрузки.!}
Ppt%5C34928-slozhnye_belki_ch1_55.jpg" alt=">Myoglobin and hemoglobin oxygen saturation curves">!}
Ppt%5C34928-slozhnye_belki_ch1_56.jpg" alt=">CO2 and H+, formed during the catabolism of organic substances, reduce the affinity of hemoglobin for O2 in proportion"> CO2 и Н+, образующиеся при катаболизме органических веществ, уменьшают сродство гемоглобина к О2 пропорционально их концентрации. Энергия, необходимая для работы клеток, вырабатывается преимущественно в митохондриях при окислении органических веществ с использованием O2, доставляемого из легких гемоглобином. В результате окисления органических веществ образуются конечные продукты их распада: СО2 и Н2O, количество которых пропорционально интенсивности протекающих процессов окисления. СO2 диффузией попадает из клеток в кровь и проникает в эритроциты, где под действием фермента карбоангидразы превращается в угольную кислоту. Эта слабая кислота диссоциирует на протон и бикарбонат ион. СО2 + Н2О → Н2СО3 → Н+ + НСО3-!}
Ppt%5C34928-slozhnye_belki_ch1_57.jpg" alt=">H+ ions are able to join His146 radicals in the β-chains of hemoglobin, i.e. in areas remote"> Ионы Н+ способны присоединятся к радикалам Гис146 в β-цепях гемоглобина, т.е. в участках, удаленных от гема. Протонирование гемоглобина снижает его сродство к О2, способствует отщеплению О2 от оксиНв, образованию дезоксиНв и увеличивает поступление кислорода в ткани пропорционально количеству образовавшихся протонов. Увеличение количества освобожденного кислорода в зависимости от увеличения концентрации Н+ в эритроцитах называется эффектом Бора (по имени датского физиолога Христиана Бора, впервые открывшего этот эффект). В легких высокое парциальное давление кислорода способствует его связыванию с дезоксиНв, что уменьшает сродство белка к Н+. Освободившиеся протоны под действием карбоангидразы взаимодействуют с бикарбонатами с образованием СО2 и Н2О!}
Ppt%5C34928-slozhnye_belki_ch1_58.jpg" alt=">Dependence of the affinity of Hb for O2 on the concentration of CO2 and protons (Bohr effect): A -"> Зависимость сродства Нв к О2 от концентрации СО2 и протонов (эффект Бора): А - влияние концентрации СО2 и Н+ на высвобождение О2 из комплекса с Нв (эффект Бора); Б - оксигенирование дезоксигемоглобина в легких, образование и выделение СО2.!}
Ppt%5C34928-slozhnye_belki_ch1_59.jpg" alt=">The resulting CO2 enters the alveolar space and is removed with exhaled air. Thus, the amount"> Образовавшийся СО2 поступает в альвеолярное пространство и удаляется с выдыхаемым воздухом. Таким образом, количество высвобождаемого гемоглобином кислорода в тканях регулируется продуктами катаболизма органических веществ: чем интенсивнее распад веществ, например при физических нагрузках, тем выше концентрация СО2 и Н+ и тем больше кислорода получают ткани в результате уменьшения сродства Нв к О2.!}
Ppt%5C34928-slozhnye_belki_ch1_60.jpg" alt=">A change in the functional activity of a protein when interacting with other ligands due to conformational changes is called allosteric"> Изменение функциональной активности белка при взаимодействии с другими лигандами вследствие конформационных изменений называется аллостерической регуляцией, а соединения-регуляторы - аллостерическими лигандами или эффекторами. Способность к аллостерической регуляции характерна, как правило, для олигомерных белков, т.е. для проявления аллостерического эффекта необходимо взаимодействие протомеров. При воздействии аллостерических лигандов белки меняют свою конформацию (в том числе и активного центра) и функцию.!}
Ppt%5C34928-slozhnye_belki_ch1_61.jpg" alt=">Allosteric regulation of the affinity of Hb for O2 by the ligand - 2,3-bis-phosphoglycerate. In erythrocytes from the product"> Аллостерическая регуляция сродства Нв к О2 лигандом - 2,3-бис-фосфоглицератом. В эритроцитах из продукта окисления глюкозы - 1,3-бисфосфоглицерата синтезируется аллостерический лиганд гемоглобина - 2,3-бисфосфоглицерат (2,3-БФГ). В нормальных условиях концентрация 2,3-БФГ высокая и сравнима с концентрацией Нв. 2,3-БФГ имеет сильный отрицательный заряд (-5).!}
Ppt%5C34928-slozhnye_belki_ch1_62.jpg" alt=">There is a cavity in the center of the tetrameric hemoglobin molecule. It is formed by amino acid residues of all four protomers."> В центре тетрамерной молекулы гемоглобина находится полость. Ее образуют аминокислотные остатки всех четырех протомеров. В капиллярах тканей протонирование Нв (эффект Бора) приводит к разрыву связи между железом гема и О2. В молекуле дезоксигемоглобина по сравнению с оксигемоглобином возникают дополнительные ионные связи, соединяющие протомеры, вследствие чего размеры центральной полости по сравнению с оксигемоглобином увеличиваются. Центральная полость является местом присоединения 2,3-БФГ к гемоглобину. БФГ поступает в полость дезоксигемоглобина. 2,3-БФГ взаимодействует с гемоглобином в участке, удаленном от активных центров белка и относится к аллостерическим (регуляторным) лигандам, а центральная полость Нв является аллостерическим центром. 2,3-БФГ имеет сильный отрицательный заряд и взаимодействует с положительно заряженными группами двух β-цепей Нв. При этом его сродство к О2 снижается в 26 раз. В результате происходит высвобождение кислорода в капиллярах ткани при низком парциальном давлении О2. В легких высокое парциальное давление О2, наоборот, приводит к оксигенированию Нв и освобождению БФГ.!}
Ppt%5C34928-slozhnye_belki_ch1_63.jpg" alt=">The BPG binding site is located in the positively charged cavity between the 4 hemoglobin protomers. BPG interaction"> Центр связывания БФГ находится в положительно заряженной полости между 4 протомерами гемоглобина. Взаимодействие БФГ с центром связывания изменяет конформацию α- и β-протомеров НЬ и их активных центров. Сродство НЬ к молекулам О2 снижается и кислород высвобождается в ткани. В легких при высоком парциальном давлении О2 активные центры гемоглобина насыщаются за счет изменения конформации и БФГ вытесняется из аллостерического центра!}
Ppt%5C34928-slozhnye_belki_ch1_64.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_65.jpg" alt=">Thus, oligomeric proteins have new properties compared to monomeric proteins. Attachment of ligands"> Таким образом, олигомерные белки обладают новыми по сравнению с мономерными белками свойствами. Присоединение лигандов на участках, пространственно удаленных друг от друга (аллостерических), способно вызывать конформационные изменения во всей белковой молекуле. Благодаря взаимодействию с регуляторными лигандами происходит изменение конформации и адаптация функции белковой молекулы к изменениям окружающей среды.!}
Ppt%5C34928-slozhnye_belki_ch1_66.jpg" alt=">About 15% of the carbon dioxide present in the blood is carried by hemoglobin molecules. In the tissues, some of the molecules"> Около 15% углекислого газа, присутствующего в крови, переносится молекулами гемоглобина. В тканях часть молекул углекислого газа может присоединится к каждому протомеру молекулы гемоглобина, при этом снижается сродство Hb к кислороду. В легких, наоборот, из-за высокого парциального давления кислорода, О2 связывается с Hb, а СО2 высвобождается.!}
Ppt%5C34928-slozhnye_belki_ch1_67.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_68.jpg" alt=">In the hemoglobin S molecule (the so-called abnormal hemoglobin), 2 β-chains turned out to be mutant, in which"> В молекуле гемоглобина S (так назван аномальный гемоглобин) мутантными оказались 2 β-цепи, в которых глутамат, высокополярная отрицательно заряженная аминокислота в положении 6 была заменена валином, содержащим гидрофобный радикал.!}
Or radicals associated with the center. atom of a complex compound. They might. ions (H - , Hal - , NO 3 - , NCS - etc.), inorg. molecules (H 2, C n, N 2, P n, O 2, Sn, CO, CO 2, NH 3, NO, SO 2, NO 2, COS, etc.), org. compounds containing elements of the main subgroups V, VI, VII gr. periodic systems or p-donor function. A large group of L.-biologically important compounds. (, peptides, purines, corrins,) and their synthetics. analogues (crown ethers, ), as well as with donor atoms and chelating groups. L. can be connected to the center. atom with s-, p- and d-two-center or multicenter bonds. In the case of the formation of two-center bonds in aluminum, donor centers can be identified (usually N, O, S, Cl, or atoms). Multicenter binding is carried out due to the aromatic p-system. L. (, cyclopentadienide anion) or heteroaromatic. L. (, thiophene, methylpyridines). The most important quantity. a characteristic of the donor-acceptor ability of a ligament is dentation, determined by the number of donor centers of a ligament involved in coordination. On this basis, L. are divided into mono-, di-, ... polydentate. Coord. the number of the complexing agent for monodentate ligaments coincides with their number, for others it is equal to the product of the number of ligaments and their dentacy. The nature of L. determines the types of coordination. conn. (, ammino complexes, mol. adducts, chelates, p-complexes, etc.); properties, structure and reaction depend on it. ability of complex connections and the possibility of their practical applications. Lit.: Garnevsky A. D., "Izvestia. Higher educational institutions, series. Chemistry and chemical technology." 1987, vol. 30, v. 10, p. 3-16; "Coord.", 1988, vol. 14, c. 5, p. 579-99; Comprehensive coordination chemistry, v. 2-Ligands, Oxf., 1987. A. D. Garnovsky.
Chemical encyclopedia. - M.: Soviet Encyclopedia. Ed. I. L. Knunyants. 1988 .
See what “LIGANDS” are in other dictionaries:
- (from the Latin ligo I bind) in complex compounds, molecules or ions associated with the central atom (complexing agent), for example. in the connection LIGATURE (Late Lat. ligatura connection) 1) a letter or sign of phonetic transcription, formed from ... ... Big Encyclopedic Dictionary
ligands- (addends) – ions, radicals or neutral molecules that are located around the central ion (atom) as a result of the formation of a coordination bond. General chemistry: textbook / A. V. Zholnin ... Chemical terms
- (from the Latin ligo I bind), in complex compounds there are molecules or ions associated with a central atom (complexing agent), for example, in the Cl3 compound there is a central Co atom, and the ligands are NH3 molecules. * * * LIGANDS LIGANDS (from Latin ligo ... encyclopedic Dictionary
- (from Latin ligo I bind) in complex compounds (See Complex compounds) molecules or ions directly associated with the central atom; same as Addends... Big Soviet encyclopedia
- (from Lat. ligo I connect), in complex containers. molecules or ions associated with a center, atom (complexing agent), e.g. in connection [Co(NH3)6]C13 center. Co atom, and L. NH3 molecule... Natural science. encyclopedic Dictionary
- ... Wikipedia
Ligands- (Latin ligo – I bind). Specific endogenous substances that excite certain types of cellular brain receptors - adrenergic, dopaminergic, cholinergic, serotonergic, benzodiazepine, peptidergic, etc. Explanatory dictionary of psychiatric terms
ligands- Andy leagues, ov, units. h.and, and... Russian spelling dictionary
Ligands- (lat. ligo bind) endogenous substances that are neurotransmitters in the synapses of the brain (dopamine, acetylcholine, serotonin, etc.). Psychotropic drugs and hallucinogens also have some ligand properties. * * * Biologically… … Encyclopedic Dictionary of Psychology and Pedagogy
axial ligands- ašiniai ligandai statusas T sritis chemija apibrėžtis Vienoje ašyje esantys ligandai. atitikmenys: engl. axial ligands rus. axial ligands ryšiai: sinonimas – aksialiniai ligandai ... Chemijos terminų aiškinamasis žodynas
Books
- Immunological problems of apoptosis, A. Yu. Baryshnikov, Yu. V. Shishkin. Last decade was marked by rapid study of the process of programmed cell death (apoptosis). Cell surface receptors and their ligands were discovered that mediate...
Module structure | Themes |
Modular unit 1 | 1.1. Structural organization of proteins. Stages of formation of the native conformation of proteins 1.2. Basics of protein functioning. Drugs as ligands affecting protein function 1.3. Denaturation of proteins and the possibility of their spontaneous renativation |
Modular unit 2 | 1.4. Features of the structure and functioning of oligomeric proteins using the example of hemoglobin 1.5. Maintenance of native protein conformation under cellular conditions 1.6. Variety of proteins. Protein families using the example of immunoglobulins 1.7. Physicochemical properties of proteins and methods for their separation |
Modular unit 1 STRUCTURAL ORGANIZATION OF MONOMERIC PROTEINS AND THE BASICS OF THEIR FUNCTIONING
Learning objectives Be able to:
1. Use knowledge about the structural features of proteins and the dependence of protein functions on their structure to understand the mechanisms of development of hereditary and acquired proteinopathies.
2. Explain the mechanisms of the therapeutic action of some drugs as ligands that interact with proteins and change their activity.
3. Use knowledge about the structure and conformational lability of proteins to understand their structural and functional instability and tendency to denaturation under changing conditions.
4. Explain the use of denaturing agents as means for sterilizing medical materials and instruments, as well as as antiseptics.
Know:
1. Levels of structural organization of proteins.
2. The importance of the primary structure of proteins, which determines their structural and functional diversity.
3. The mechanism of formation of the active center in proteins and its specific interaction with the ligand, which underlies the functioning of proteins.
4. Examples of the influence of exogenous ligands (drugs, toxins, poisons) on the conformation and functional activity of proteins.
5. Causes and consequences of protein denaturation, factors causing denaturation.
6. Examples of the use of denaturing factors in medicine as antiseptics and means for sterilizing medical instruments.
TOPIC 1.1. STRUCTURAL ORGANIZATION OF PROTEINS. STAGES OF FORMATION OF NATIVE
PROTEIN CONFORMATIONS
Proteins are polymer molecules whose monomers are only 20 α-amino acids. The set and order of combination of amino acids in a protein is determined by the structure of genes in the DNA of individuals. Each protein, in accordance with its specific structure, performs its own function. The set of proteins of a given organism determines its phenotypic characteristics, as well as the presence of hereditary diseases or a predisposition to their development.
1. Amino acids that make up proteins. Peptide bond. Proteins are polymers built from monomers - 20 α-amino acids, the general formula of which is
Amino acids differ in structure, size, and physicochemical properties of radicals attached to the α-carbon atom. The functional groups of amino acids determine the characteristics of the properties of different α-amino acids. The radicals found in α-amino acids can be divided into several groups:
Proline, Unlike the other 19 protein monomers, it is not an amino acid, but an imino acid; the radical in proline is associated with both the α-carbon atom and the imino group
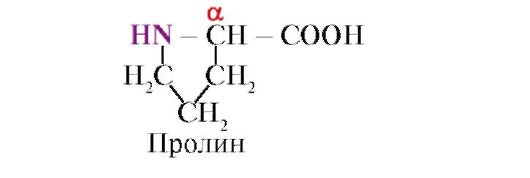 Amino acids vary in solubility in water. This is due to the ability of radicals to interact with water (hydrate).
Amino acids vary in solubility in water. This is due to the ability of radicals to interact with water (hydrate).
TO hydrophilic include radicals containing anionic, cationic and polar uncharged functional groups.
TO hydrophobic include radicals containing methyl groups, aliphatic chains or rings.
2. Peptide bonds connect amino acids to form peptides. During peptide synthesis, the α-carboxyl group of one amino acid interacts with the α-amino group of another amino acid to form peptide bond:
Proteins are polypeptides, i.e. linear polymers of α-amino acids connected by a peptide bond (Fig. 1.1.)
 Rice. 1.1. Terms used to describe the structure of peptides
Rice. 1.1. Terms used to describe the structure of peptides
The monomers of amino acids that make up polypeptides are called amino acid residues. A chain of repeating groups - NH-CH-CO- forms peptide backbone. An amino acid residue that has a free α-amino group is called N-terminal, and one that has a free α-carboxyl group is called C-terminal. Peptides are written and read from N-terminus to C-terminus.
The peptide bond formed by the imino group of proline differs from other peptide bonds: the nitrogen atom of the peptide group lacks a hydrogen,
instead, there is a bond with a radical, as a result of which one side of the ring is included in the peptide backbone:
 Peptides differ in amino acid composition, number of amino acids and order of amino acid connection, for example, Ser-Ala-Glu-Gis and His-Glu-Ala-Ser are two different peptides.
Peptides differ in amino acid composition, number of amino acids and order of amino acid connection, for example, Ser-Ala-Glu-Gis and His-Glu-Ala-Ser are two different peptides.
Peptide bonds are very strong, and their chemical non-enzymatic hydrolysis requires harsh conditions: the protein being analyzed is hydrolyzed in concentrated hydrochloric acid at a temperature of about 110° for 24 hours. In a living cell, peptide bonds can be broken by proteolytic enzymes, called proteases or peptide hydrolases.
3. Primary structure of proteins. Amino acid residues in the peptide chains of different proteins do not alternate randomly, but are arranged in a certain order. The linear sequence or alternation order of amino acid residues in a polypeptide chain is called primary structure of the protein.
The primary structure of each individual protein is encoded in the DNA molecule (in a region called the gene) and is realized during transcription (copying information onto mRNA) and translation (synthesis of the primary structure of the protein). Consequently, the primary structure of the proteins of an individual person is information hereditarily transmitted from parents to children, which determines the structural features of the proteins of a given organism, on which the function of the existing proteins depends (Fig. 1.2.).
 Rice. 1.2. The relationship between the genotype and the conformation of proteins synthesized in the individual’s body
Rice. 1.2. The relationship between the genotype and the conformation of proteins synthesized in the individual’s body
Each of the approximately 100,000 individual proteins in the human body has unique primary structure. Molecules of the same type of protein (for example, albumin) have the same alternation of amino acid residues, which distinguishes albumin from any other individual protein.
The sequence of amino acid residues in a peptide chain can be considered a form of recording information. This information determines the spatial arrangement of the linear peptide chain into a more compact three-dimensional structure called conformation squirrel. The process of formation of a functionally active protein conformation is called folding
4. Protein conformation. Free rotation in the peptide backbone is possible between the nitrogen atom of the peptide group and the neighboring α-carbon atom, as well as between the α-carbon atom and the carbon of the carbonyl group. Due to the interaction of functional groups of amino acid residues, the primary structure of proteins can acquire more complex spatial structures. In globular proteins, there are two main levels of folding of the conformation of peptide chains: secondary And tertiary structure.
Secondary structure of proteins is a spatial structure formed as a result of the formation of hydrogen bonds between the functional groups -C=O and -NH- of the peptide backbone. In this case, the peptide chain can acquire regular structures of two types: α-helices And β-structures.
IN α-helices hydrogen bonds are formed between the oxygen atom of the carbonyl group and the hydrogen of the amide nitrogen of the 4th amino acid from it; side chains of amino acid residues
are located along the periphery of the spiral, without participating in the formation of the secondary structure (Fig. 1.3.).
Bulk radicals, or radicals carrying equal charges, prevent the formation of an α-helix. The proline residue, which has a ring structure, interrupts the α-helix, since due to the lack of hydrogen at the nitrogen atom in the peptide chain it is impossible to form a hydrogen bond. The bond between nitrogen and the α-carbon atom is part of the proline ring, so the peptide backbone becomes bent at this point.
β-Structure is formed between the linear regions of the peptide backbone of one polypeptide chain, thereby forming folded structures. Polypeptide chains or parts thereof can form parallel or antiparallel β-structures. In the first case, the N- and C-termini of the interacting peptide chains coincide, and in the second they have the opposite direction (Fig. 1.4).
Rice. 1.3. Protein secondary structure - α-helix
 Rice. 1.4. Parallel and antiparallel β-sheet structures
Rice. 1.4. Parallel and antiparallel β-sheet structures
β-structures are indicated by wide arrows: A - Antiparallel β-structure. B - Parallel β-sheet structures
In some proteins, β-structures can be formed due to the formation of hydrogen bonds between atoms of the peptide backbone of different polypeptide chains.
Also found in proteins areas with irregular secondary structure, which includes bends, loops, and turns of the polypeptide backbone. They are often located in places where the direction of the peptide chain changes, for example, when a parallel β-sheet structure is formed.
Based on the presence of α-helices and β-structures, globular proteins can be divided into four categories.
Rice. 1.5. Secondary structure of myoglobin (A) and hemoglobin β-chain (B), containing eight α-helices

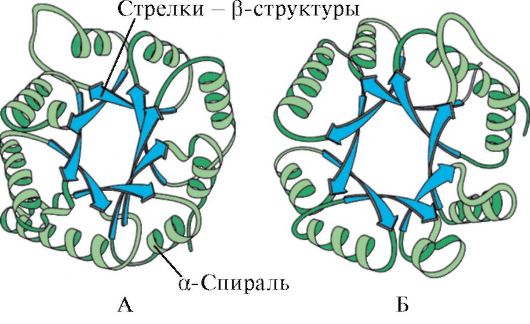 Rice. 1.6. Secondary structure of triosephosphate isomerase and pyruvate kinase domain
Rice. 1.6. Secondary structure of triosephosphate isomerase and pyruvate kinase domain
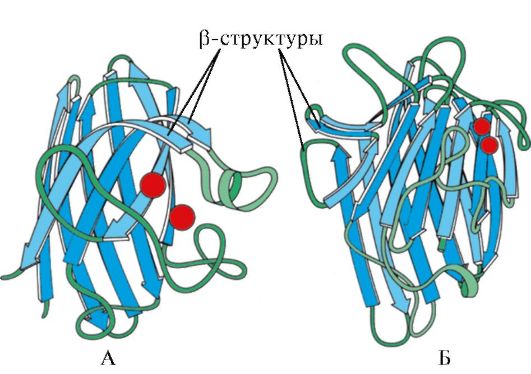 Rice. 1.7. Secondary structure of the constant domain of immunoglobulin (A) and the enzyme superoxide dismutase (B)
Rice. 1.7. Secondary structure of the constant domain of immunoglobulin (A) and the enzyme superoxide dismutase (B)
IN fourth category included proteins that contain a small amount of regular secondary structures. These proteins include small cysteine-rich proteins or metalloproteins.
Protein tertiary structure- a type of conformation formed due to interactions between amino acid radicals, which can be located at a considerable distance from each other in the peptide chain. Most proteins form a spatial structure resembling a globule (globular proteins).
Since hydrophobic amino acid radicals tend to combine through so-called hydrophobic interactions and intermolecular van der Waals forces, a dense hydrophobic core is formed inside the protein globule. Hydrophilic ionized and non-ionized radicals are mainly located on the surface of the protein and determine its solubility in water.
 Rice. 1.8. Types of bonds that arise between amino acid radicals during the formation of the tertiary structure of a protein
Rice. 1.8. Types of bonds that arise between amino acid radicals during the formation of the tertiary structure of a protein
1 - ionic bond- occurs between positively and negatively charged functional groups;
2 - hydrogen bond- occurs between a hydrophilic uncharged group and any other hydrophilic group;
3 - hydrophobic interactions- arise between hydrophobic radicals;
4 - disulfide bond- formed due to the oxidation of SH groups of cysteine residues and their interaction with each other
Hydrophilic amino acid residues located inside the hydrophobic core can interact with each other using ionic And hydrogen bonds(Fig. 1.8).
Ionic and hydrogen bonds, as well as hydrophobic interactions, are weak: their energy is not much higher than the energy of thermal motion of molecules at room temperature. The conformation of the protein is maintained by the formation of many such weak bonds. Since the atoms that make up a protein are in constant motion, it is possible to break some weak bonds and form others, which leads to slight movements of individual sections of the polypeptide chain. This property of proteins to change conformation as a result of breaking some and forming other weak bonds is called conformational lability.
The human body has systems that support homeostasis- constancy of the internal environment within certain acceptable limits for a healthy body. Under homeostasis conditions, small changes in conformation do not disrupt the overall structure and function of proteins. The functionally active conformation of a protein is called native conformation. Changes in the internal environment (for example, the concentration of glucose, Ca ions, protons, etc.) lead to changes in the conformation and disruption of protein functions.
The tertiary structure of some proteins is stabilized disulfide bonds, formed due to the interaction of -SH groups of two residues
 Rice. 1.9. Formation of a disulfide bond in a protein molecule
Rice. 1.9. Formation of a disulfide bond in a protein molecule
cysteine (Fig. 1.9). Most intracellular proteins do not have covalent disulfide bonds in their tertiary structure. Their presence is characteristic of proteins secreted by the cell, which ensures their greater stability in extracellular conditions. Thus, disulfide bonds are present in the molecules of insulin and immunoglobulins.
Insulin- a protein hormone synthesized in the β-cells of the pancreas and secreted into the blood in response to an increase in the concentration of glucose in the blood. In the structure of insulin, there are two disulfide bonds connecting the polypeptide A and B chains, and one disulfide bond within the A chain (Fig. 1.10).
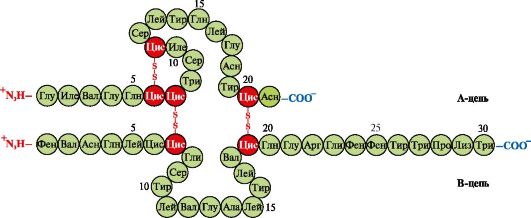 Rice. 1.10. Disulfide bonds in the structure of insulin
Rice. 1.10. Disulfide bonds in the structure of insulin
5. Supersecondary structure of proteins. In proteins with different primary structure and functions, they are sometimes detected similar combinations and relative positions of secondary structures, which are called supersecondary structure. It occupies an intermediate position between the secondary and tertiary structures, since it is a specific combination of elements of the secondary structure in the formation of the tertiary structure of the protein. Supersecondary structures have specific names, such as “α-helix-turn-a-helix,” “leucine zipper,” “zinc fingers,” etc. Such supersecondary structures are characteristic of DNA-binding proteins.
"Leucine zipper." This type of supersecondary structure is used to join two proteins together. On the surface of interacting proteins there are α-helical regions containing at least four leucine residues. Leucine residues in the α-helix are located six amino acids apart. Since each turn of the α-helix contains 3.6 amino acid residues, leucine radicals are located on the surface of every second turn. Leucine residues of the α-helix of one protein can interact with leucine residues of another protein (hydrophobic interactions), connecting them together (Fig. 1.11.). Many DNA binding proteins function in oligomeric complexes where the individual subunits are linked to each other by “leucine zippers.”
 Rice. 1.11. "Leucine zipper" between α-helical regions of two proteins
Rice. 1.11. "Leucine zipper" between α-helical regions of two proteins
An example of such proteins are histones. Histones- nuclear proteins, which contain a large number of positively charged amino acids - arginine and lysine (up to 80%). Histone molecules are combined into oligomeric complexes containing eight monomers using “leucine zippers”, despite the significant homonymous charge of these molecules.
"Zinc finger"- a variant of supersecondary structure, characteristic of DNA-binding proteins, has the form of an elongated fragment on the surface of the protein and contains about 20 amino acid residues (Fig. 1.12). The “extended finger” shape is supported by a zinc atom bound to four amino acid radicals - two cysteine residues and two histidine residues. In some cases, instead of histidine residues, there are cysteine residues. Two closely lying cysteine residues are separated from the other two Gisili residues by a Cys sequence consisting of approximately 12 amino acid residues. This region of the protein forms an α-helix, the radicals of which can specifically bind to the regulatory regions of the major groove of DNA. Individual binding specificity
 Rice. 1.12. The primary structure of the region of DNA-binding proteins that form the “zinc finger” structure (the letters indicate the amino acids that make up this structure)
Rice. 1.12. The primary structure of the region of DNA-binding proteins that form the “zinc finger” structure (the letters indicate the amino acids that make up this structure)
The regulatory DNA binding protein depends on the sequence of amino acid residues located in the zinc finger region. Such structures contain, in particular, receptors for steroid hormones involved in the regulation of transcription (reading information from DNA to RNA).
TOPIC 1.2. BASICS OF PROTEIN FUNCTIONING. DRUGS AS LIGANDS AFFECTING PROTEIN FUNCTION
1. The active center of the protein and its interaction with the ligand. During the formation of the tertiary structure, a region is formed on the surface of a functionally active protein, usually in a recess, formed by amino acid radicals that are far apart from each other in the primary structure. This region, which has a unique structure for a given protein and is capable of specifically interacting with a particular molecule or group of similar molecules, is called the protein-ligand binding site or active site. Ligands are molecules that interact with proteins.
High specificity The interaction of the protein with the ligand is ensured by the complementarity of the structure of the active center to the structure of the ligand.
Complementarity- this is the spatial and chemical correspondence of interacting surfaces. The active center must not only spatially correspond to the ligand included in it, but also bonds (ionic, hydrogen, and hydrophobic interactions) must be formed between the functional groups of the radicals included in the active center and the ligand, which hold the ligand in the active center (Fig. 1.13 ).
 Rice. 1.13. Complementary interaction of protein with ligand
Rice. 1.13. Complementary interaction of protein with ligand
Some ligands, when attached to the active center of a protein, play an auxiliary role in the functioning of proteins. Such ligands are called cofactors, and proteins containing a non-protein part are called complex proteins(as opposed to simple proteins, consisting only of the protein part). The non-protein part, firmly connected to the protein, is called prosthetic group. For example, myoglobin, hemoglobin and cytochromes contain a prosthetic group, heme, containing an iron ion, firmly attached to the active center. Complex proteins containing heme are called hemoproteins.
When specific ligands are attached to proteins, the function of these proteins is manifested. Thus, albumin, the most important protein in blood plasma, exhibits its transport function by attaching hydrophobic ligands, such as fatty acids, bilirubin, some drugs, etc. to the active center (Fig. 1.14)
Ligands interacting with the three-dimensional structure of the peptide chain can be not only low-molecular organic and inorganic molecules, but also macromolecules:
DNA (examples with DNA-binding proteins discussed above);
Polysaccharides;
 Rice. 1.14. Relationship between genotype and phenotype
Rice. 1.14. Relationship between genotype and phenotype
The unique primary structure of human proteins, encoded in the DNA molecule, is realized in cells in the form of a unique conformation, active center structure and protein functions
In these cases, the protein recognizes a specific region of the ligand that is commensurate and complementary to the binding site. Thus, on the surface of hepatocytes there are receptor proteins for the hormone insulin, which also has a protein structure. The interaction of insulin with the receptor causes a change in its conformation and activation of signaling systems, leading to the storage of nutrients in hepatocytes after meals.
Thus, The functioning of proteins is based on the specific interaction of the active center of the protein with the ligand.
2. Domain structure and its role in the functioning of proteins. Long polypeptide chains of globular proteins often fold into several compact, relatively independent regions. They have an independent tertiary structure, reminiscent of that of globular proteins, and are called domains. Due to the domain structure of proteins, their tertiary structure is easier to form.
In domain proteins, ligand binding sites are often located between domains. Thus, trypsin is a proteolytic enzyme that is produced by the exocrine part of the pancreas and is necessary for the digestion of food proteins. It has a two-domain structure, and the center of binding of trypsin with its ligand - food protein - is located in the groove between the two domains. In the active center, the conditions necessary for effective binding of a specific site of food protein and hydrolysis of its peptide bonds are created.
Different domains in a protein can move relative to each other when the active center interacts with the ligand (Fig. 1.15).
Hexokinase- an enzyme that catalyzes the phosphorylation of glucose using ATP. The active site of the enzyme is located in the cleft between the two domains. When hexokinase binds to glucose, the domains surrounding it close and the substrate becomes trapped, where phosphorylation occurs (see Fig. 1.15).
 Rice. 1.15. Binding of hexokinase domains to glucose
Rice. 1.15. Binding of hexokinase domains to glucose
In some proteins, domains perform independent functions by binding to various ligands. Such proteins are called multifunctional.
3. Drugs are ligands that affect the function of proteins. The interaction of proteins with ligands is specific. However, due to the conformational lability of the protein and its active center, it is possible to select another substance that could also interact with the protein in the active center or other part of the molecule.
A substance similar in structure to a natural ligand is called structural analogue of the ligand or a non-natural ligand. It also interacts with the protein at the active site. A structural analogue of a ligand can both enhance protein function (agonist), and reduce it (antagonist). The ligand and its structural analogues compete with each other for binding to the protein at the same site. Such substances are called competitive modulators(regulators) of protein functions. Many drugs act as protein inhibitors. Some of them are obtained by chemical modification of natural ligands. Inhibitors of protein functions can be drugs and poisons.
Atropine is a competitive inhibitor of M-cholinergic receptors. Acetylcholine is a neurotransmitter for the transmission of nerve impulses through cholinergic synapses. To carry out excitation, acetylcholine released into the synaptic cleft must interact with the receptor protein of the postsynaptic membrane. Two types found cholinergic receptors:
M receptor in addition to acetylcholine, it selectively interacts with muscarine (fly agaric toxin). M - cholinergic receptors are present on smooth muscles and, when interacting with acetylcholine, cause their contraction;
H receptor specifically binding to nicotine. N-cholinergic receptors are found at the synapses of striated skeletal muscles.
Specific inhibitor M-cholinergic receptors is atropine. It is found in belladonna and henbane plants.
 Atropine has functional groups similar in structure to acetylcholine and their spatial arrangement, therefore it is a competitive inhibitor of M-cholinergic receptors. Considering that the binding of acetylcholine to M-cholinergic receptors causes contraction of smooth muscles, atropine is used as a medicine that relieves their spasm (antispasmodic). Thus, it is known to use atropine to relax the eye muscles when viewing the fundus, as well as to relieve spasms during gastrointestinal colic. M-cholinergic receptors are also present in the central nervous system(CNS), therefore large doses of atropine can cause an undesirable reaction from the central nervous system: motor and mental agitation, hallucinations, convulsions.
Atropine has functional groups similar in structure to acetylcholine and their spatial arrangement, therefore it is a competitive inhibitor of M-cholinergic receptors. Considering that the binding of acetylcholine to M-cholinergic receptors causes contraction of smooth muscles, atropine is used as a medicine that relieves their spasm (antispasmodic). Thus, it is known to use atropine to relax the eye muscles when viewing the fundus, as well as to relieve spasms during gastrointestinal colic. M-cholinergic receptors are also present in the central nervous system(CNS), therefore large doses of atropine can cause an undesirable reaction from the central nervous system: motor and mental agitation, hallucinations, convulsions.
Ditilin is a competitive agonist of H-cholinergic receptors, inhibiting the function of neuromuscular synapses.
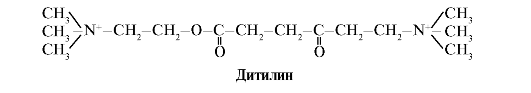 Neuromuscular synapses of skeletal muscles contain H-cholinergic receptors. Their interaction with acetylcholine leads to muscle contractions. During some surgical operations, as well as in endoscopic studies, drugs are used that cause relaxation of skeletal muscles (muscle relaxants). These include dithiline, which is a structural analogue of acetylcholine. It attaches to H-cholinergic receptors, but unlike acetylcholine, it is very slowly destroyed by the enzyme acetylcholinesterase. As a result of prolonged opening of ion channels and persistent depolarization of the membrane, the conduction of nerve impulses is disrupted and muscle relaxation occurs. Initially, these properties were discovered in curare poison, which is why such drugs are called curare-like.
Neuromuscular synapses of skeletal muscles contain H-cholinergic receptors. Their interaction with acetylcholine leads to muscle contractions. During some surgical operations, as well as in endoscopic studies, drugs are used that cause relaxation of skeletal muscles (muscle relaxants). These include dithiline, which is a structural analogue of acetylcholine. It attaches to H-cholinergic receptors, but unlike acetylcholine, it is very slowly destroyed by the enzyme acetylcholinesterase. As a result of prolonged opening of ion channels and persistent depolarization of the membrane, the conduction of nerve impulses is disrupted and muscle relaxation occurs. Initially, these properties were discovered in curare poison, which is why such drugs are called curare-like.
TOPIC 1.3. DENATURATION OF PROTEINS AND THE POSSIBILITY OF THEIR SPONTANEOUS RENATIVATION
1. Since the native conformation of proteins is maintained due to weak interactions, changes in the composition and properties of the environment surrounding the protein, exposure to chemical reagents and physical factors cause a change in their conformation (the property of conformational lability). Breaking a large number of bonds leads to the destruction of the native conformation and denaturation of proteins.
Denaturation of proteins- this is the destruction of their native conformation under the influence of denaturing agents, caused by the rupture of weak bonds that stabilize the spatial structure of the protein. Denaturation is accompanied by the destruction of the unique three-dimensional structure and active center of the protein and the loss of its biological activity (Fig. 1.16).
All denatured molecules of one protein acquire a random conformation that differs from other molecules of the same protein. The amino acid radicals that form the active center turn out to be spatially distant from each other, i.e. the specific binding site of the protein with the ligand is destroyed. During denaturation, the primary structure of proteins remains unchanged.
Application of denaturing agents in biological research and medicine. In biochemical studies, before determining low molecular weight compounds in biological material, proteins are usually first removed from the solution. Trichloroacetic acid (TCA) is most often used for this purpose. After adding TCA to the solution, denatured proteins precipitate and are easily removed by filtration (Table 1.1.)
In medicine, denaturing agents are often used to sterilize medical instruments and materials in autoclaves (denaturing agent is high temperature) and as antiseptics (alcohol, phenol, chloramine) for treating contaminated surfaces containing pathogenic microflora.
2. Spontaneous protein reactivation- proof of the determinism of the primary structure, conformation and function of proteins. Individual proteins are products of one gene that have an identical amino acid sequence and acquire the same conformation in the cell. The fundamental conclusion that the primary structure of a protein already contains information about its conformation and function was made on the basis of the ability of some proteins (in particular, ribonuclease and myoglobin) to spontaneously renativate - restore their native conformation after denaturation.
The formation of spatial protein structures is carried out by the method of self-assembly - a spontaneous process in which a polypeptide chain, which has a unique primary structure, tends to adopt a conformation with the lowest free energy in solution. The ability to renativate proteins that retain their primary structure after denaturation was described in an experiment with the enzyme ribonuclease.
Ribonuclease is an enzyme that breaks down the bonds between individual nucleotides in an RNA molecule. This globular protein has one polypeptide chain, the tertiary structure of which is stabilized by many weak and four disulfide bonds.
Treatment of ribonuclease with urea, which breaks hydrogen bonds in the molecule, and a reducing agent, which breaks disulfide bonds, leads to denaturation of the enzyme and loss of its activity.
Removal of denaturing agents by dialysis leads to restoration of protein conformation and function, i.e. to rebirth. (Fig. 1.17).
 Rice. 1.17. Denaturation and renativation of ribonuclease
Rice. 1.17. Denaturation and renativation of ribonuclease
A - native conformation of ribonuclease, in the tertiary structure of which there are four disulfide bonds; B - denatured ribonuclease molecule;
B - reactivated ribonuclease molecule with restored structure and function
1. Fill out table 1.2.
Table 1.2. Classification of amino acids according to the polarity of radicals
2. Write the formula of the tetrapeptide:
Asp - Pro - Fen - Liz
a) highlight the repeating groups in the peptide that form the peptide backbone and the variable groups represented by amino acid radicals;
b) label the N- and C-termini;
c) highlight the peptide bonds;
d) write another peptide consisting of the same amino acids;
d) count the quantity possible options tetrapeptide with a similar amino acid composition.
3. Explain the role of the primary structure of proteins using the example of a comparative analysis of two structurally similar and evolutionarily close peptide hormones of the mammalian neurohypophysis - oxytocin and vasopressin (Table 1.3).
Table 1.3. Structure and functions of oxytocin and vasopressin
 For this:
For this:
a) compare the composition and amino acid sequence of two peptides;
b) find the similarity of the primary structure of the two peptides and the similarity of their biological action;
c) find differences in the structure of two peptides and differences in their functions;
d) draw a conclusion about the influence of the primary structure of peptides on their functions.
4. Describe the main stages of formation of the conformation of globular proteins (secondary, tertiary structures, the concept of supersecondary structure). Indicate the types of bonds involved in the formation of protein structures. Which amino acid radicals can participate in the formation of hydrophobic interactions, ionic, hydrogen bonds.
Give examples.
5. Define the concept of “conformational lability of proteins”, indicate the reasons for its existence and significance.
6. Expand the meaning of the following phrase: “The functioning of proteins is based on their specific interaction with the ligand,” using the terms and explaining their meaning: protein conformation, active center, ligand, complementarity, protein function.
7. Using one example, explain what domains are and what their role is in the functioning of proteins.
SELF-CONTROL TASKS
1. Match.
Functional group in amino acid radical:
A. Carboxyl group B. Hydroxyl group C Guanidine group D. Thiol group E. Amino group
2. Choose the correct answers.
Amino acids with polar uncharged radicals are:
A. Cis B. Asn
B. Glu G. Three
3. Choose the correct answers.
Amino acid radicals:
A. Provide specificity of the primary structure B. Participate in the formation of the tertiary structure
B. Located on the surface of the protein, they influence its solubility D. Form the active center
D. Participate in the formation of peptide bonds
4. Choose the correct answers.
Hydrophobic interactions can form between amino acid radicals:
A. Tre Lay B. Pro Three
B. Met Ile G. Tir Ala D. Val Fen
5. Choose the correct answers.
Ionic bonds can form between amino acid radicals:
A. Gln Asp B. Apr Liz
B. Liz Glu G. Gis Asp D. Asn Apr
6. Choose the correct answers.
Hydrogen bonds can form between amino acid radicals:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Match.
Type of bond involved in the formation of protein structure:
A. Primary structure B. Secondary structure
B. Tertiary structure
D. Supersecondary structure E. Conformation.
1. Hydrogen bonds between atoms of the peptide backbone
2. Weak bonds between functional groups of amino acid radicals
3. Bonds between α-amino and α-carboxyl groups of amino acids
8. Choose the correct answers. Trypsin:
A. Proteolytic enzyme B. Contains two domains
B. Hydrolyzes starch
D. The active site is located between the domains. D. Consists of two polypeptide chains.
9. Choose the correct answers. Atropine:
A. Neurotransmitter
B. Structural analogue of acetylcholine
B. Interacts with H-cholinergic receptors
D. Strengthens the conduction of nerve impulses through cholinergic synapses
D. Competitive inhibitor of M-cholinergic receptors
10. Choose the correct statements. In proteins:
A. The primary structure contains information about the structure of its active site
B. The active center is formed at the level of the primary structure
B. The conformation is rigidly fixed by covalent bonds
D. The active site can interact with a group of similar ligands
due to the conformational lability of proteins D. Changes in the environment can affect the affinity of the active
center to ligand
1. 1-B, 2-G, 3-B.
3. A, B, C, D.
7. 1-B, 2-D, 3-A.
8. A, B, C, D.
BASIC TERMS AND CONCEPTS
1. Protein, polypeptide, amino acids
2. Primary, secondary, tertiary protein structures
3. Conformation, native protein conformation
4. Covalent and weak bonds in protein
5. Conformational lability
6. Protein active site
7. Ligands
8. Protein folding
9. Structural analogues of ligands
10. Domain proteins
11. Simple and complex proteins
12. Protein denaturation, denaturing agents
13. Reactivation of proteins
Solve problems
“Structural organization of proteins and the basis of their functioning”
1. The main function of the protein - hemoglobin A (HbA) is the transport of oxygen to tissues. In the human population, multiple forms of this protein are known with altered properties and function - the so-called abnormal hemoglobins. For example, hemoglobin S, found in the red blood cells of patients with sickle cell disease (HbS), has been found to have low solubility under conditions of low partial pressure of oxygen (as is the case in venous blood). This leads to the formation of aggregates of this protein. The protein loses its function, precipitates, and the red blood cells become irregularly shaped (some of them form a sickle shape) and are destroyed faster than usual in the spleen. As a result, sickle cell anemia develops.
The only difference in the primary structure of HbA was found in the N-terminal region of the hemoglobin β-chain. Compare the N-terminal regions of the β-strand and show how changes in the primary structure of the protein affect its properties and functions.
 For this:
For this:
a) write the formulas of amino acids by which HbA differ and compare the properties of these amino acids (polarity, charge).
b) draw a conclusion about the reason for the decrease in solubility and disruption of oxygen transport into tissues.
2. The figure shows a diagram of the structure of a protein that has a binding center with a ligand (active center). Explain why the protein is selective in its choice of ligand. For this:
a) remember what the active center of a protein is and consider the structure of the active center of the protein shown in the figure;
b) write the formulas of the amino acid radicals that make up the active center;
c) draw a ligand that could specifically interact with the active site of the protein. Indicate on it the functional groups that can form bonds with the amino acid radicals that make up the active center;
d) indicate the types of bonds that arise between the ligand and amino acid radicals of the active center;
e) explain what the specificity of protein-ligand interaction is based on.
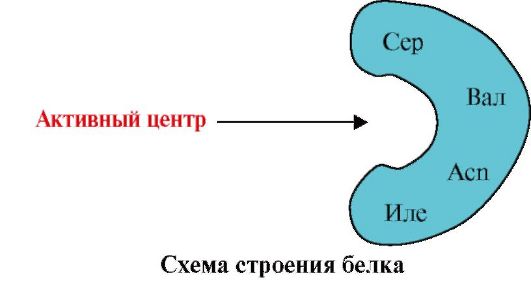 3.
The figure shows the active site of the protein and several ligands.
3.
The figure shows the active site of the protein and several ligands.
Determine which ligand is most likely to interact with the active site of the protein and why.
 What types of bonds arise during the formation of a protein-ligand complex?
What types of bonds arise during the formation of a protein-ligand complex?
4. Structural analogues of natural protein ligands can be used as drugs to modify the activity of proteins.
Acetylcholine is a mediator of excitation transmission at neuromuscular synapses. When acetylcholine interacts with proteins - receptors of the postsynaptic membrane of skeletal muscles, ion channels open and muscle contraction occurs. Ditilin is a medicine used in some operations to relax muscles, as it disrupts the transmission of nerve impulses through neuromuscular synapses. Explain the mechanism of action of ditilin as a muscle relaxant. For this:
a) write the formulas of acetylcholine and dithiline and compare their structures;
b) describe the mechanism of the relaxing effect of ditilin.
5. In some diseases, the patient's body temperature rises, which is considered as a protective reaction of the body. However, high temperatures are detrimental to body proteins. Explain why at temperatures above 40 °C the function of proteins is disrupted and a threat to human life arises. To do this, remember:
1) The structure of proteins and the bonds that hold its structure in the native conformation;
2) How does the structure and function of proteins change with increasing temperature?;
3) What is homeostasis and why is it important for maintaining human health.
Modular unit 2 OLIGOMERICK PROTEINS AS TARGETS OF REGULATORY INFLUENCES. STRUCTURAL AND FUNCTIONAL DIVERSITY OF PROTEINS. METHODS FOR SEPARATION AND PURIFICATION OF PROTEINS
Learning objectives Be able to:
1. Use knowledge about the features of the structure and functions of oligomeric proteins to understand the adaptive mechanisms of regulation of their functions.
2. Explain the role of chaperones in the synthesis and maintenance of protein conformation under cellular conditions.
3. Explain the variety of manifestations of life by the variety of structures and functions of proteins synthesized in the body.
4. Analyze the relationship between the structure of proteins and their function using examples of comparison of related hemoproteins - myoglobin and hemoglobin, as well as representatives of the five classes of proteins of the immunoglobulin family.
5. Apply knowledge about the peculiarities of the physical and chemical properties of proteins to select methods for their purification from other proteins and impurities.
6. Interpret the results of quantitative and quality composition blood plasma proteins to confirm or clarify the clinical diagnosis.
Know:
1. Features of the structure of oligomeric proteins and adaptive mechanisms for regulating their functions using the example of hemoglobin.
2. The structure and functions of chaperones and their importance for maintaining the native conformation of proteins under cellular conditions.
3. Principles of combining proteins into families based on the similarity of their conformation and functions using the example of immunoglobulins.
4. Methods for separating proteins based on the characteristics of their physicochemical properties.
5. Electrophoresis of blood plasma as a method for assessing the qualitative and quantitative composition of proteins.
TOPIC 1.4. FEATURES OF THE STRUCTURE AND FUNCTIONING OF OLIGOMERIC PROTEINS USING THE EXAMPLE OF HEMOGLOBIN
1. Many proteins contain several polypeptide chains. Such proteins are called oligomeric, and individual chains - protomers. Protomers in oligomeric proteins are connected by many weak non-covalent bonds (hydrophobic, ionic, hydrogen). Interaction
protomers are carried out thanks to complementarity their contacting surfaces.
The number of protomers in oligomeric proteins can vary greatly: hemoglobin contains 4 protomers, the enzyme aspartate aminotransferase has 12 protomers, and the tobacco mosaic virus protein contains 2120 protomers connected by non-covalent bonds. Consequently, oligomeric proteins can have very high molecular weights.
The interaction of one protomer with others can be considered a special case of protein-ligand interaction, since each protomer serves as a ligand for other protomers. The number and method of joining protomers in a protein is called quaternary protein structure.
Proteins can contain protomers of the same or different structures, for example, homodimers are proteins containing two identical protomers, and heterodimers are proteins containing two different protomers.
If proteins contain different protomers, then binding centers with different ligands that differ in structure can be formed on them. When a ligand binds to the active site, the function of this protein is manifested. A center located on a different protomer is called allosteric (different from the active one). Contacting allosteric ligand or effector, it performs a regulatory function (Fig. 1.18). The interaction of the allosteric center with the effector causes conformational changes in the structure of the entire oligomeric protein due to its conformational lability. This affects the affinity of the active site for a specific ligand and regulates the function of that protein. A change in the conformation and function of all protomers during the interaction of an oligomeric protein with at least one ligand is called cooperative conformational changes. Effectors that enhance protein function are called activators, and effectors that inhibit its function - inhibitors.
Thus, oligomeric proteins, as well as proteins with a domain structure, have a new property compared to monomeric proteins - the ability to allosterically regulate functions (regulation by attaching different ligands to the protein). This can be seen by comparing the structures and functions of two closely related complex proteins, myoglobin and hemoglobin.
 Rice. 1.18. Scheme of the structure of a dimeric protein
Rice. 1.18. Scheme of the structure of a dimeric protein
2. Formation of spatial structures and functioning of myoglobin.
Myoglobin (Mb) is a protein found in red muscles, the main function of which is to create O 2 reserves necessary for intense muscle work. Mb is a complex protein containing a protein part - apoMb and a non-protein part - heme. The primary structure of apoMB determines its compact globular conformation and the structure of the active center, to which the non-protein part of myoglobin, heme, is attached. Oxygen coming from the blood to the muscles binds to Fe+ 2 hemes in myoglobin. Mb is a monomeric protein that has a very high affinity for O 2, therefore the release of oxygen by myoglobin occurs only during intense muscular work, when the partial pressure of O 2 sharply decreases.
Formation of the Mv conformation. In red muscles, on ribosomes during translation, the primary structure of MB is synthesized, represented by a specific sequence of 153 amino acid residues. The secondary structure of Mb contains eight α-helices, called in Latin letters from A to H, between which there are non-helical regions. The tertiary structure of Mb has the form of a compact globule, in the recess of which the active center is located between the F and E α-helices (Fig. 1.19).
 Rice. 1.19. Myoglobin structure
Rice. 1.19. Myoglobin structure
3. Features of the structure and functioning of the active center of MV. The active center of Mb is formed predominantly by hydrophobic amino acid radicals, widely spaced from each other in the primary structure (for example, Tri 3 9 and Fen 138) Poorly soluble in water ligands - heme and O 2 - attach to the active center. Heme is a specific ligand of apoMB (Fig. 1.20), the basis of which is made up of four pyrrole rings connected by methenyl bridges; in the center there is a Fe+ 2 atom connected to the nitrogen atoms of the pyrrole rings by four coordination bonds. In the active center of Mb, in addition to hydrophobic amino acid radicals, there are also residues of two amino acids with hydrophilic radicals - Gis E 7(Gis 64) and GIS F 8(His 93) (Fig. 1.21).
 Rice. 1.20. The structure of heme - the non-protein part of myoglobin and hemoglobin
Rice. 1.20. The structure of heme - the non-protein part of myoglobin and hemoglobin
 Rice. 1.21. Location of heme and O2 in the active site of apomyoglobin and hemoglobin protomers
Rice. 1.21. Location of heme and O2 in the active site of apomyoglobin and hemoglobin protomers
Heme is covalently bound to His F8 through an iron atom. O 2 attaches to the iron on the other side of the heme plane. His E 7 is necessary for the correct orientation of O 2 and facilitates the addition of oxygen to Fe + 2 heme
GIS F 8 forms a coordination bond with Fe+ 2 and firmly fixes heme in the active center. Gis E 7 necessary for correct orientation in the active center of another ligand - O 2 during its interaction with Fe + 2 heme. The microenvironment of heme creates conditions for strong but reversible binding of O 2 to Fe + 2 and prevents water from entering the hydrophobic active site, which can lead to its oxidation to Fe + 3.
The monomeric structure of Mb and its active center determines the high affinity of the protein for O 2.
4. Oligomeric structure of Hb and regulation of the affinity of Hb to O 2 ligands. Human hemoglobins- a family of proteins, like myoglobin, related to complex proteins (hemoproteins). They have a tetrameric structure and contain two α-chains, but differ in the structure of the other two polypeptide chains (2α-, 2x-chains). The structure of the second polypeptide chain determines the features of the functioning of these forms of Hb. About 98% of the hemoglobin in an adult's red blood cells is hemoglobin A(2α-, 2p-chains).
During fetal development, two main types of hemoglobins function: embryonic Hb(2α, 2ε), which is found in the early stages of fetal development, and hemoglobin F (fetal)- (2α, 2γ), which replaces early fetal hemoglobin in the sixth month of intrauterine development and only after birth is replaced by Hb A.
HB A is a protein related to myoglobin (MB) found in adult human red blood cells. The structure of its individual protomers is similar to that of myoglobin. The secondary and tertiary structures of myoglobin and hemoglobin protomers are very similar, despite the fact that in the primary structure of their polypeptide chains only 24 amino acid residues are identical (the secondary structure of hemoglobin protomers, like myoglobin, contains eight α-helices, designated by Latin letters from A to H , and the tertiary structure has the form of a compact globule). But unlike myoglobin, hemoglobin has an oligomeric structure, consisting of four polypeptide chains connected by non-covalent bonds (Figure 1.22).
Each Hb protomer is associated with a non-protein part - heme and neighboring protomers. The connection of the protein part of Hb with heme is similar to that of myoglobin: in the active center of the protein, the hydrophobic parts of the heme are surrounded by hydrophobic amino acid radicals with the exception of His F 8 and His E 7, which are located on both sides of the heme plane and play a similar role in the functioning of the protein and its binding with oxygen (see structure of myoglobin).
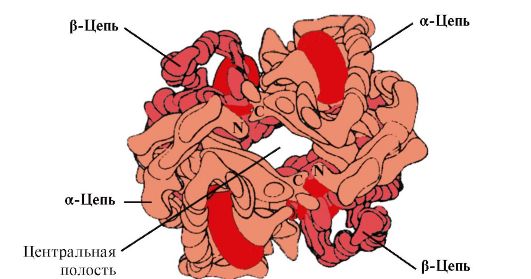 Rice. 1.22. Oligomeric structure of hemoglobin
Rice. 1.22. Oligomeric structure of hemoglobin
Besides, Gis E 7 performs an important additional role in the functioning of Nv. Free heme has a 25,000 times higher affinity for CO than for O2. CO is formed in small quantities in the body and, given its high affinity for heme, it could disrupt the transport of O 2 necessary for cell life. However, in the composition of hemoglobin, the affinity of heme for carbon monoxide exceeds the affinity for O 2 by only 200 times due to the presence of His E 7 in the active center. The remainder of this amino acid creates optimal conditions for the binding of heme to O 2 and weakens the interaction of heme with CO.
5. The main function of HB is the transport of O2 from the lungs to the tissues. Unlike monomeric myoglobin, which has a very high affinity for O2 and performs the function of storing oxygen in red muscles, the oligomeric structure of hemoglobin provides:
1) rapid saturation of HB with oxygen in the lungs;
2) the ability of HB to release oxygen in tissues at a relatively high partial pressure of O 2 (20-40 mm Hg);
3) the possibility of regulating the affinity of Hb to O 2.
6. Cooperative changes in the conformation of hemoglobin protomers accelerate the binding of O 2 in the lungs and its release into tissues. In the lungs, the high partial pressure of O 2 promotes its binding to Hb in the active site of four protomers (2α and 2β). The active center of each protomer, as in myoglobin, is located between two α-helices (F and E) in a hydrophobic pocket. It contains a non-protein part - heme, attached to the protein part by many weak hydrophobic interactions and one strong bond between Fe 2 + heme and His F 8 (see Fig. 1.21).
In deoxyhemoglobin, due to this bond with His F 8, the Fe 2 + atom protrudes from the heme plane towards histidine. The binding of O 2 to Fe 2 + occurs on the other side of the heme in the His E 7 region using a single free coordination bond. His E 7 provides optimal conditions for the binding of O 2 to heme iron.
The addition of O 2 to the Fe + 2 atom of one protomer causes its movement into the heme plane, followed by the histidine residue associated with it
 Rice. 1.23. Change in the conformation of the hemoglobin protomer when combined with O 2
Rice. 1.23. Change in the conformation of the hemoglobin protomer when combined with O 2
This leads to a change in the conformation of all polypeptide chains due to their conformational lability. Changing the conformation of other chains facilitates their interaction with subsequent O 2 molecules.
The fourth O 2 molecule attaches to hemoglobin 300 times easier than the first (Fig. 1.24).
 Rice. 1.24. Cooperative changes in the conformation of hemoglobin protomers during its interaction with O2
Rice. 1.24. Cooperative changes in the conformation of hemoglobin protomers during its interaction with O2
In tissues, each subsequent O 2 molecule is cleaved off more easily than the previous one, also due to cooperative changes in the conformation of protomers.
7. CO 2 and H+, formed during the catabolism of organic substances, reduce the affinity of hemoglobin for O 2 in proportion to their concentration. The energy required for cell functioning is produced primarily in mitochondria during the oxidation of organic substances using O 2 delivered from the lungs by hemoglobin. As a result of the oxidation of organic substances, the final products of their decomposition are formed: CO 2 and K 2 O, the amount of which is proportional to the intensity of the ongoing oxidation processes.
CO 2 diffuses from cells into the blood and penetrates into red blood cells, where, under the action of the enzyme carbanhydrase, it is converted into carbonic acid. This weak acid dissociates into a proton and a bicarbonate ion.
H+ are capable of joining His radicals 14 6 in the α- and β-chains of hemoglobin, i.e. in areas distant from the heme. Protonation of hemoglobin reduces its affinity for O 2, promotes the removal of O 2 from oxyHb, the formation of deoxyHb, and increases the supply of oxygen to tissues in proportion to the number of protons formed (Fig. 1.25).
An increase in the amount of released oxygen depending on the increase in H+ concentration in red blood cells is called the Bohr effect (named after the Danish physiologist Christian Bohr, who first discovered this effect).
In the lungs, a high partial pressure of oxygen promotes its binding to deoxyHb, which reduces the protein's affinity for H +. The released protons under the action of carbonic acid react with bicarbonates to form CO 2 and H 2 O
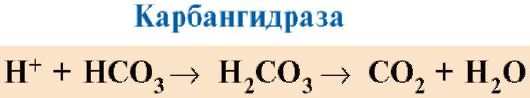
 Rice. 1.25. Dependence of the affinity of Hb for O 2 on the concentration of CO 2 and protons (Bohr effect):
Rice. 1.25. Dependence of the affinity of Hb for O 2 on the concentration of CO 2 and protons (Bohr effect):
A- influence of CO 2 and H+ concentration on the release of O 2 from the complex with HB (Bohr effect); B- oxygenation of deoxyhemoglobin in the lungs, formation and release of CO 2.
The resulting CO 2 enters the alveolar space and is removed with exhaled air. Thus, the amount of oxygen released by hemoglobin in tissues is regulated by the products of catabolism of organic substances: the more intense the breakdown of substances, for example during physical exercise, the higher the concentration of CO 2 and H + and the more oxygen the tissues receive as a result of a decrease in the affinity of Hb for O 2.
8. Allosteric regulation of the affinity of Hb for O2 by the ligand - 2,3-bisphosphoglycerate. In erythrocytes, the allosteric ligand of hemoglobin, 2,3-bisphosphoglycerate (2,3-BPG), is synthesized from the product of glucose oxidation - 1,3-bisphosphoglycerate. Under normal conditions, the concentration of 2,3-BPG is high and comparable to the concentration of Hb. 2,3-BPG has a strong negative charge of -5.
 Bisphosphoglycerate in tissue capillaries, binding to deoxyhemoglobin, increases the release of oxygen into tissues, reducing the affinity of Hb for O 2.
Bisphosphoglycerate in tissue capillaries, binding to deoxyhemoglobin, increases the release of oxygen into tissues, reducing the affinity of Hb for O 2.
At the center of the tetrameric hemoglobin molecule is a cavity. It is formed by amino acid residues of all four protomers (see Fig. 1.22). In tissue capillaries, protonation of Hb (Bohr effect) leads to the rupture of the bond between heme iron and O2. In a molecule
deoxyhemoglobin, compared to oxyhemoglobin, additional ionic bonds appear that connect the protomers, as a result of which the dimensions of the central cavity increase compared to oxyhemoglobin. The central cavity is the site of attachment of 2,3-BPG to hemoglobin. Due to the difference in the size of the central cavity, 2,3-BPG can only attach to deoxyhemoglobin.
2,3-BPG interacts with hemoglobin in a site remote from the active centers of the protein and belongs to allosteric(regulatory) ligands, and the central cavity of Hb is allosteric center. 2,3-BPG has a strong negative charge and interacts with five positively charged groups of the two β-chains of Hb: the N-terminal α-amino group of Val and the Lys 82 His 143 radicals (Fig. 1.26).
 Rice. 1.26. BPG in the central cavity of deoxyhemoglobin
Rice. 1.26. BPG in the central cavity of deoxyhemoglobin
BPG binds to three positively charged groups on each β-strand.
In tissue capillaries, the resulting deoxyhemoglobin interacts with 2,3-BPG and ionic bonds are formed between the positively charged radicals of the β-chains and the negatively charged ligand, which change the conformation of the protein and reduce the affinity of Hb for O2. A decrease in the affinity of Hb for O 2 contributes to a more efficient release of O 2 into the tissue.
In the lungs, at high partial pressure, oxygen interacts with Hb, joining heme iron; in this case, the conformation of the protein changes, the central cavity decreases and 2,3-BPG is displaced from the allosteric center
Thus, oligomeric proteins have new properties compared to monomeric proteins. Attachment of ligands at sites
spatially distant from each other (allosteric), can cause conformational changes in the entire protein molecule. Due to interaction with regulatory ligands, a change in conformation and adaptation of the function of the protein molecule to environmental changes occurs.
TOPIC 1.5. MAINTENANCE OF NATIVE CONFORMATION OF PROTEINS UNDER CELL CONDITIONS
In cells, during the synthesis of polypeptide chains, their transport through membranes to the corresponding parts of the cell, during the process of folding (formation of the native conformation) and during the assembly of oligomeric proteins, as well as during their functioning, intermediate, aggregation-prone, unstable conformations arise in the protein structure. Hydrophobic radicals, usually hidden inside the protein molecule in the native conformation, appear on the surface in an unstable conformation and tend to combine with groups of other proteins that are poorly soluble in water. In everyone's cells known organisms Special proteins have been discovered that ensure optimal folding of cell proteins, stabilize their native conformation during functioning and, most importantly, maintain the structure and functions of intracellular proteins in the event of disturbances in homeostasis. These proteins are called "chaperones" which means “nanny” in French.
1. Molecular chaperones and their role in preventing protein denaturation.
Chaperones (CH) are classified according to the mass of their subunits. High molecular weight chaperones have a mass from 60 to 110 kDa. Among them, three classes have been most studied: Sh-60, Sh-70 and Sh-90. Each class includes a family of related proteins. Thus, Sh-70 includes proteins with a molecular weight from 66 to 78 kDa. Low molecular weight chaperones have a molecular weight from 40 to 15 kDa.
Among the chaperones there are constitutive proteins, the high basal synthesis of which does not depend on stress effects on the body’s cells, and inducible, the synthesis of which under normal conditions is weak, but increases sharply under stress. Inducible chaperones are also called “heat shock proteins” because they were first discovered in cells exposed to high temperatures. In cells, due to the high concentration of proteins, spontaneous reactivation of partially denatured proteins is difficult. Sh-70 can prevent the onset of denaturation and help restore the native conformation of proteins. Molecular chaperones-70- a highly conserved class of proteins found in all parts of the cell: cytoplasm, nucleus, endoplasmic reticulum, mitochondria. At the carboxyl end of the single polypeptide chain Ш-70 there is a region that is a groove capable of interacting with peptides of length
from 7 to 9 amino acid residues enriched with hydrophobic radicals. Such regions in globular proteins occur approximately every 16 amino acids. Sh-70 is capable of protecting proteins from temperature inactivation and restoring the conformation and activity of partially denatured proteins.
2. The role of chaperones in protein folding. During protein synthesis on the ribosome, the N-terminal region of the polypeptide is synthesized before the C-terminal one. To form the native conformation, the complete amino acid sequence of the protein is required. In the process of protein synthesis, chaperones-70, due to the structure of their active center, are able to close areas of the polypeptide that are prone to aggregation, enriched in hydrophobic amino acid radicals until the synthesis is completed (Figure 1.27, A).
 Rice. 1.27. Participation of chaperones in protein folding
Rice. 1.27. Participation of chaperones in protein folding
A - participation of chaperones-70 in the prevention of hydrophobic interactions between sections of the synthesized polypeptide; B - formation of the native conformation of the protein in the chaperone complex
Many high-molecular proteins that have a complex conformation, such as a domain structure, fold in a special space formed by Sh-60. Ш-60 function as an oligomeric complex consisting of 14 subunits. They form two hollow rings, each of which consists of seven subunits, these rings are connected to each other. Each Sh-60 subunit consists of three domains: apical (apical), enriched with hydrophobic radicals facing the cavity of the ring, intermediate and equatorial (Fig. 1.28).
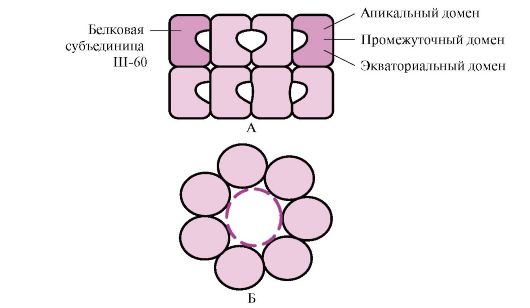 Rice. 1.28. Structure of the chaperonin complex consisting of 14 Ш-60
Rice. 1.28. Structure of the chaperonin complex consisting of 14 Ш-60
A - side view; B - top view
Synthesized proteins, which have elements on the surface characteristic of unfolded molecules, in particular hydrophobic radicals, enter the cavity of the chaperone rings. In the specific environment of these cavities, possible conformations are searched until the only one that is energetically most favorable is found (Fig. 1.27, B). The formation of conformations and protein release is accompanied by ATP hydrolysis in the equatorial region. Typically, such chaperone-dependent folding requires a significant amount of energy.
In addition to participating in the formation of the three-dimensional structure of proteins and the renativation of partially denatured proteins, chaperones are also necessary for the occurrence of such fundamental processes as the assembly of oligomeric proteins, recognition and transport of denatured proteins into lysosomes, transport of proteins across membranes, and participation in the regulation of the activity of protein complexes.
TOPIC 1.6. VARIETY OF PROTEINS. PROTEIN FAMILIES: EXAMPLE OF IMMUNOGLOBULINS
1. Proteins play a decisive role in the life of individual cells and the entire multicellular organism, and their functions are surprisingly diverse. This is determined by the characteristics of the primary structure and conformations of proteins, the unique structure of the active center and the ability to bind specific ligands.
Only a very small fraction of all possible variants of peptide chains can adopt a stable spatial structure; majority
of them can take many conformations with approximately the same Gibbs energy, but with different properties. The primary structure of most known proteins selected by biological evolution ensures exceptional stability of one of the conformations, which determines the characteristics of the functioning of this protein.
2. Protein families. Within one biological species substitutions of amino acid residues can lead to the emergence of different proteins that perform related functions and have homologous amino acid sequences. Such related proteins have strikingly similar conformations: the number and relative positions of α-helices and/or β-structures, and most of the turns and bends of the polypeptide chains are similar or identical. Proteins with homologous regions of the polypeptide chain, similar conformation and related functions are classified into protein families. Examples of protein families: serine proteinases, immunoglobulin family, myoglobin family.
Serine proteinases- a family of proteins that perform the function of proteolytic enzymes. These include digestive enzymes - chymotrypsin, trypsin, elastase and many blood clotting factors. These proteins have identical amino acids in 40% of their positions and a very similar conformation (Fig. 1.29).

Rice. 1.29. Spatial structures of elastase (A) and chymotrypsin (B)
Some amino acid substitutions have led to changes in the substrate specificity of these proteins and the emergence of functional diversity within the family.
3. Immunoglobulin family. In the functioning of the immune system, proteins of the immunoglobulin superfamily play a huge role, which includes three families of proteins:
Antibodies (immunoglobulins);
T-lymphocyte receptors;
Proteins of the major histocompatibility complex - MHC classes 1 and 2 (Major Histocompatibility Complex).
All these proteins have a domain structure, consist of homologous immune-like domains and perform similar functions: they interact with foreign structures, either dissolved in the blood, lymph or intercellular fluid (antibodies), or located on the surface of cells (own or foreign).
4. Antibodies- specific proteins produced by B lymphocytes in response to the entry of a foreign structure into the body, called antigen.
Features of the structure of antibodies
The simplest antibody molecules consist of four polypeptide chains: two identical light ones - L, containing about 220 amino acids, and two identical heavy ones - H, consisting of 440-700 amino acids. All four chains in the antibody molecule are connected by many non-covalent bonds and four disulfide bonds (Fig. 1.30).
Antibody light chains consist of two domains: a variable domain (VL), located in the N-terminal region of the polypeptide chain, and a constant domain (CL), located at the C-terminus. Heavy chains usually have four domains: one variable (VH), located at the N-terminus, and three constant domains (CH1, CH2, CH3) (see Fig. 1.30). Each immunoglobulin domain has a β-sheet superstructure in which two cysteine residues are linked by a disulfide bond.
Between the two constant domains CH1 and CH2 there is a region containing a large number of proline residues, which prevent the formation of a secondary structure and the interaction of neighboring H-chains in this segment. This hinge region gives the antibody molecule flexibility. Between the variable domains of the heavy and light chains there are two identical antigen-binding sites (active sites for binding antigens), therefore such antibodies are often called bivalents. Not the entire amino acid sequence of the variable regions of both chains is involved in the binding of the antigen to the antibody, but only 20-30 amino acids located in the hypervariable regions of each chain. It is these regions that determine the unique ability of each type of antibody to interact with the corresponding complementary antigen.
Antibodies are one of the body's lines of defense against invading foreign organisms. Their functioning can be divided into two stages: the first stage is recognition and binding of antigen on the surface of foreign organisms, which is possible due to the presence of antigen-binding sites in the antibody structure; the second stage is the initiation of the process of inactivation and destruction of the antigen. The specificity of the second stage depends on the class of antibodies. There are five classes of heavy chains, differing from each other in the structure of constant domains: α, δ, ε, γ and μ, according to which five classes of immunoglobulins are distinguished: A, D, E, G and M.
The structural features of heavy chains give the hinge regions and C-terminal regions of heavy chains a conformation characteristic of each class. After antigen binds to an antibody, conformational changes in the constant domains determine the pathway for antigen removal.
 Rice. 1. 30. Domain structure of IgG
Rice. 1. 30. Domain structure of IgG
Immunoglobulins M
Immunoglobulins M have two forms.
Monomeric form- 1st class of antibodies produced by developing B lymphocytes. Subsequently, many B cells switch to producing other classes of antibodies, but with the same antigen-binding site. IgM is embedded in the membrane and acts as an antigen recognition receptor. The integration of IgM into the cell membrane is possible due to the presence of 25 hydrophobic amino acid residues in the tail part of the region.
Secretory form of IgM contains five monomeric subunits linked to each other by disulfide bonds and an additional polypeptide J chain (Fig. 1.31). The heavy chains of monomers of this form do not contain a hydrophobic tail. The pentamer has 10 antigen binding sites and is therefore effective in recognizing and removing the antigen that first enters the body. The secretory form of IgM is the main class of antibodies secreted into the blood during the primary immune response. The binding of IgM to an antigen changes the conformation of IgM and induces its binding to the first protein component of the complement system (the complement system is a set of proteins involved in the destruction of the antigen) and activation of this system. If the antigen is located on the surface of a microorganism, the complement system causes disruption of the integrity of the cell membrane and death of the bacterial cell.
Immunoglobulins G
Quantitatively, this class of immunoglobulins predominates in the blood (75% of all Igs). IgG - monomers, the main class of antibodies secreted into the blood during a secondary immune response. After the interaction of IgG with the surface antigens of microorganisms, the antigen-antibody complex is able to bind and activate proteins of the complement system or can interact with specific receptors of macrophages and neutrophils. Interaction with phagocytes leads
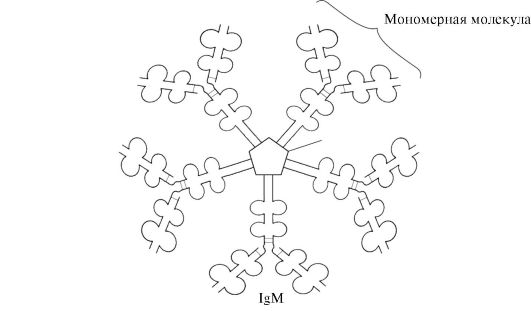 Rice. 1.31. Structure of the secretory form of IgM
Rice. 1.31. Structure of the secretory form of IgM
to the absorption of antigen-antibody complexes and their destruction in cell phagosomes. IgG is the only class of antibodies that are able to penetrate the placental barrier and provide intrauterine protection of the fetus from infections.
Immunoglobulins A
The main class of antibodies present in secretions (milk, saliva, secretions of the respiratory tract and intestinal tract). IgA is secreted primarily in dimeric form, where the monomers are linked to each other through an additional J chain (Fig. 1.32).
IgA does not interact with the complement system and phagocytic cells, but by binding to microorganisms, antibodies prevent their attachment to epithelial cells and penetration into the body.
Immunoglobulins E
Immunoglobulins E are represented by monomers in which the heavy ε-chains contain, like the μ-chains of immunoglobulins M, one variable and four constant domains. After secretion, IgE binds with its
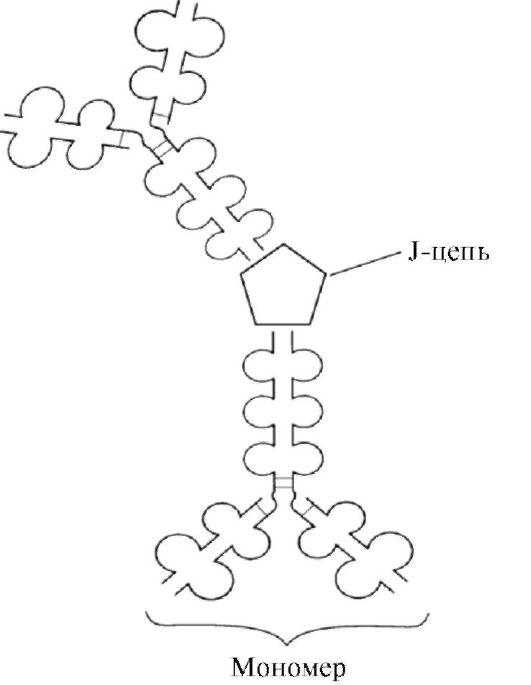 Rice. 1.32. Structure of IgA
Rice. 1.32. Structure of IgA
C-terminal regions with corresponding receptors on the surface of mast cells and basophils. As a result, they become receptors for antigens on the surface of these cells (Fig. 1.33).
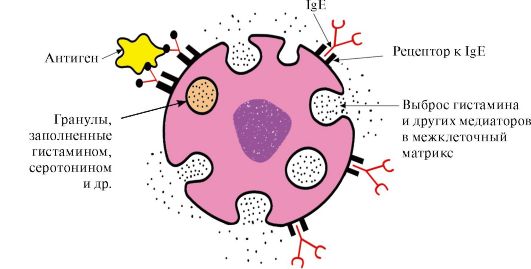 Rice. 1.33. Interaction of IgE with antigen on the surface of a mast cell
Rice. 1.33. Interaction of IgE with antigen on the surface of a mast cell
After the antigen attaches to the corresponding antigen-binding sites of IgE, the cells receive a signal to secrete biologically active substances (histamine, serotonin), which are largely responsible for the development of the inflammatory reaction and for the manifestation of allergic reactions such as asthma, urticaria, hay fever.
Immunoglobulins D
Immunoglobulins D are found in very small quantities in serum; they are monomers. Heavy δ chains have one variable and three constant domains. IgDs act as receptors for B lymphocytes; other functions are still unknown. The interaction of specific antigens with receptors on the surface of B-lymphocytes (IgD) leads to the transmission of these signals into the cell and the activation of mechanisms that ensure the proliferation of a given lymphocyte clone.
TOPIC 1.7. PHYSICAL AND CHEMICAL PROPERTIES OF PROTEINS AND METHODS FOR THEIR SEPARATION
1. Individual proteins differ in physical and chemical properties:
Shape of molecules;
Molecular weight;
The total charge, the magnitude of which depends on the ratio of anionic and cationic groups of amino acids;
The ratio of polar and non-polar amino acid radicals on the surface of molecules;
Degrees of resistance to various denaturing agents.
2. Protein solubility depends on the properties of the proteins listed above, as well as on the composition of the medium in which the protein is dissolved (pH values, salt composition, temperature, the presence of other organic substances that can interact with the protein). The amount of charge of protein molecules is one of the factors affecting their solubility. When the charge at the isoelectric point is lost, proteins more easily aggregate and precipitate. This is especially typical for denatured proteins, in which hydrophobic amino acid radicals appear on the surface.
On the surface of a protein molecule there are both positively and negatively charged amino acid radicals. The number of these groups, and therefore the total charge of proteins, depends on the pH of the medium, i.e. ratio of concentrations of H+ - and OH - groups. In an acidic environment An increase in H+ concentration leads to suppression of the dissociation of carboxyl groups -COO - + H+ > - COOH and a decrease in the negative charge of proteins. In an alkaline environment, the binding of excess OH - by protons formed during the dissociation of amino groups -NH 3 + + OH - - NH 2 + H 2 O with the formation of water, leads to a decrease in the positive charge of proteins. The pH value at which a protein has a net zero charge is called isoelectric point (IEP). In IET, the number of positively and negatively charged groups is the same, i.e. the protein is in an isoelectric state.
3. Separation of individual proteins. Features of the structure and functioning of the body depend on the set of proteins synthesized in it. Studying the structure and properties of proteins is impossible without isolating them from the cell and purifying them from other proteins and organic molecules. Stages of isolation and purification of individual proteins:
Cell destruction the tissue being studied and obtaining a homogenate.
Separation of the homogenate into fractions by centrifugation, obtaining a nuclear, mitochondrial, cytosolic or other fraction containing the desired protein.
Selective thermal denaturation- short-term heating of a protein solution, during which some of the denatured protein impurities can be removed (if the protein is relatively heat-stable).
Salting out. Different proteins precipitate at different salt concentrations in solution. By gradually increasing the salt concentration, it is possible to obtain a number of separate fractions with a predominant content of isolated protein in one of them. Ammonium sulfate is most often used for protein fractionation. Proteins with the least solubility precipitate at low salt concentrations.
Gel filtration- a method of sifting molecules through swollen Sephadex granules (three-dimensional polysaccharide chains of dextran having pores). The speed at which proteins pass through a column filled with Sephadex will depend on their molecular weight: the smaller the mass of protein molecules, the easier they penetrate into the granules and stay there longer; the larger the mass, the faster they elute from the column.
Ultracentrifugation- a method that involves placing proteins in a centrifuge tube into the rotor of an ultracentrifuge. When the rotor rotates, the sedimentation rate of proteins is proportional to their molecular weight: fractions of heavier proteins are located closer to the bottom of the test tube, lighter ones - closer to the surface.
Electrophoresis- a method based on differences in the speed of movement of proteins in an electric field. This value is proportional to the charge of the proteins. Electrophoresis of proteins is carried out on paper (in this case, the speed of protein movement is proportional only to their charge) or in a polyacrylamide gel with a certain pore size (the speed of protein movement is proportional to their charge and molecular weight).
Ion exchange chromatography- a fractionation method based on the binding of ionized groups of proteins with oppositely charged groups of ion exchange resins (insoluble polymeric materials). The strength of the binding of the protein to the resin is proportional to the charge of the protein. Proteins adsorbed to the ion-exchange polymer can be washed away with increasing concentrations of NaCl solutions; the lower the protein charge, the lower the concentration of NaCl required to wash away the protein bound to the ionic groups of the resin.
Affinity chromatography- the most specific method for isolating individual proteins. A ligand of a protein is covalently attached to an inert polymer. When a protein solution is passed through a column with a polymer, only the protein specific for a given ligand is adsorbed on the column due to the complementary binding of the protein to the ligand.
Dialysis- a method used to remove low molecular weight compounds from a solution of isolated protein. The method is based on the inability of proteins to pass through a semi-permeable membrane, unlike low molecular weight substances. It is used to purify proteins from low-molecular impurities, for example, salts after salting out.
ASSIGNMENTS FOR EXTRACURRICULAR WORK
1. Fill out the table. 1.4.
Table 1.4. Comparative analysis of the structure and functions of related proteins - myoglobin and hemoglobin
a) remember the structure of the active center of Mb and Hb. What role do hydrophobic amino acid radicals play in the formation of the active centers of these proteins? Describe the structure of the active center of Mb and Hb and the mechanisms of attachment of ligands to it. What role do the His F 8 and His E 7 residues play in the functioning of the active center of Mv iHv?
b) what new properties compared to monomeric myoglobin does the closely related oligomeric protein, hemoglobin, have? Explain the role of cooperative changes in the conformation of protomers in the hemoglobin molecule, the effect of CO 2 and proton concentrations on the affinity of hemoglobin for oxygen, as well as the role of 2,3-BPG in the allosteric regulation of Hb function.
2. Characterize molecular chaperones, paying attention to the relationship between their structure and function.
3. What proteins are grouped into families? Using the example of the immunoglobulin family, identify similar structural features and related functions of the proteins of this family.
4. Purified individual proteins are often required for biochemical and medicinal purposes. Explain on what physicochemical properties of proteins the methods used for their separation and purification are based.
SELF-CONTROL TASKS
1. Choose the correct answers.
Functions of hemoglobin:
A. Transport of O 2 from the lungs to the tissue B. Transport of H + from the tissues to the lungs
B. Maintaining a constant blood pH D. Transport of CO 2 from the lungs to the tissues
D. Transport of CO 2 from tissues to lungs
2. Choose the correct answers. Ligandα -protomer Hb is: A. Heme
B. Oxygen
B. CO G. 2,3-BPG
D. β-Protomer
3. Choose the correct answers.
Hemoglobin as opposed to myoglobin:
A. Has a quaternary structure
B. The secondary structure is represented only by α-helices
B. Belongs to complex proteins
D. Interacts with an allosteric ligand D. Covalently bound to heme
4. Choose the correct answers.
The affinity of Hb for O2 decreases:
A. When one O 2 molecule is added B. When one O 2 molecule is removed
B. When interacting with 2,3-BPG
D. When attached to protomers H + D. When the concentration of 2,3-BPG decreases
5. Match.
Types HB are characterized by:
A. In deoxy form it forms fibrillar aggregates B. Contains two α- and two δ-chains
B. The predominant form of Hb in adult erythrocytes D. Contains heme with Fe+ 3 in the active center
D. Contains two α- and two γ-chains 1. HbA 2.
6. Match.
Hb ligands:
A. Binds to Hb in the allosteric center
B. Has a very high affinity for the active site of Hb
B. By joining, it increases the affinity of Hb for O 2 G. Oxidizes Fe+ 2 to Fe+ 3
D. Forms a covalent bond with hisF8
7. Choose the correct answers.
Chaperones:
A. Proteins present in all parts of the cell
B. Synthesis increases under stress
B. Participate in the hydrolysis of denatured proteins
D. Participate in maintaining the native conformation of proteins
D. They create organelles in which the conformation of proteins is formed.
8. Match. Immunoglobulins:
A. The secretory form is pentameric.
B. Class Ig that penetrates the placental barrier
B. Ig - mast cell receptor
D. The main class of Ig present in the secretions of epithelial cells. D. B-lymphocyte receptor, the activation of which ensures cell proliferation
9. Choose the correct answers.
Immunoglobulins E:
A. Produced by macrophages B. They have heavy ε chains.
B. Embedded in the membrane of T-lymphocytes
D. Act as membrane antigen receptors on mast cells and basophils
D. Responsible for allergic reactions
10. Choose the correct answers.
The method for separating proteins is based on differences in their molecular weight:
A. Gel filtration
B. Ultracentrifugation
B. Polyacrylamide gel electrophoresis D. Ion exchange chromatography
D. Affinity chromatography
11. Choose the correct answer.
The method for separating proteins is based on differences in their solubility in water:
A. Gel filtration B. Salting out
B. Ion exchange chromatography D. Affinity chromatography
D. Electrophoresis in polyacrylamide gel
STANDARDS OF ANSWERS TO “SELF-CONTROL TASKS”
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-B
BASIC TERMS AND CONCEPTS
1. Oligomeric proteins, protomer, quaternary structure of proteins
2. Cooperative changes in protomer conformation
3. Bohr effect
4. Allosteric regulation of protein functions, allosteric center and allosteric effector
5. Molecular chaperones, heat shock proteins
6. Protein families (serine proteases, immunoglobulins)
7. IgM-, G-, E-, A-structure-function relationship
8. Total charge of proteins, isoelectric point of proteins
9. Electrophoresis
10. Salting out
11. Gel filtration
12. Ion exchange chromatography
13. Ultracentrifugation
14. Affinity chromatography
15. Electrophoresis of blood plasma proteins
TASKS FOR CLASSROOM WORK
1. Compare the dependences of the degrees of saturation of hemoglobin (Hb) and myoglobin (Mb) with oxygen on its partial pressure in tissues
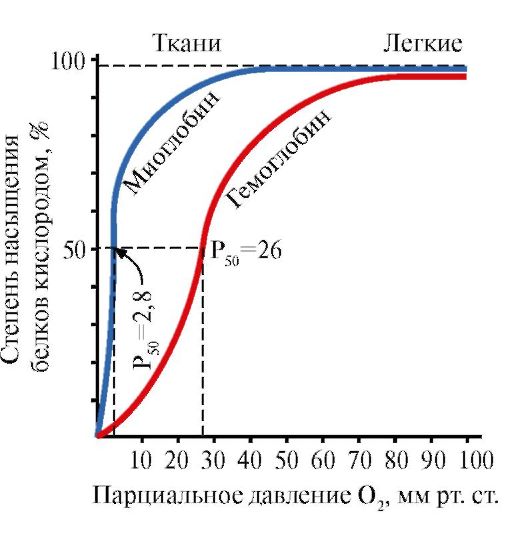 Rice. 1.34. Dependence of saturation Mv andNHoxygen from its partial pressure
Rice. 1.34. Dependence of saturation Mv andNHoxygen from its partial pressure
Please note that the shape of the protein oxygen saturation curves is different: for myoglobin - a hyperbola, for hemoglobin - a sigmoid shape.
1. compare the values of the partial pressure of oxygen at which Mb and Hb are saturated with O 2 by 50%. Which of these proteins has a higher affinity for O 2?
2. What structural features of Mb determine its high affinity for O 2?
3. What structural features of HB allow it to release O2 in the capillaries of resting tissues (at a relatively high partial pressure of O2) and sharply increase this release in working muscles? What property of oligomeric proteins provides this effect?
4. Calculate what amount of O 2 (in%) oxygenated hemoglobin gives to resting and working muscles?
5. draw conclusions about the relationship between the structure of a protein and its function.
2. The amount of oxygen released by hemoglobin in the capillaries depends on the intensity of catabolic processes in tissues (Bohr effect). How do changes in tissue metabolism regulate the affinity of Hb for O2? Effect of CO 2 and H+ on the affinity of Hb for O 2
 1. describe the Bohr effect.
1. describe the Bohr effect.
2. in what direction does the process shown in the diagram proceed:
a) in the capillaries of the lungs;
b) in tissue capillaries?
3. What is the physiological significance of the Bohr effect?
4. Why does the interaction of Hb with H+ at sites remote from the heme change the affinity of the protein for O 2?
3. The affinity of Hb for O2 depends on the concentration of its ligand - 2,3-bisphosphoglycerate, which is an allosteric regulator of the affinity of Hb for O2. Why does ligand interaction at a site distant from the active site affect protein function? How does 2,3-BPG regulate the affinity of Hb for O2? To solve the problem, answer the following questions:
1. where and from what is 2.3-bisphosphoglycerate (2,3-BPG) synthesized? Write its formula, indicate the charge of this molecule.
2. Which form of hemoglobin (oxy or deoxy) does BPG interact with and why? In what part of the Hb molecule does the interaction occur?
3. in what direction does the process shown in the diagram occur?
a) in tissue capillaries;
b) in the capillaries of the lungs?
4. where the concentration of the complex should be higher
Nv-2,3-BFG:
a) in the capillaries of muscles at rest,
b) in the capillaries of working muscles (provided the same concentration of BPG in erythrocytes)?
5. How will the affinity of HB for oxygen change when a person adapts to high altitude conditions, if the concentration of BPG in erythrocytes increases? What is the physiological significance of this phenomenon?
4. The destruction of 2,3-BPG during storage of preserved blood impairs the functions of HB. How will the affinity of HB for O 2 change in preserved blood if the concentration of 2,3-BPG in erythrocytes can decrease from 8 to 0.5 mmol/l. Is it possible to transfuse such blood to seriously ill patients if the concentration of 2,3-BPG is restored no earlier than after three days? Is it possible to restore the functions of red blood cells by adding 2,3-BPG to the blood?
5. Remember the structure of the simplest immunoglobulin molecules. What role do immunoglobulins play in the immune system? Why are Igs often called bivalents? How does the structure of Igs relate to their function? (Describe using an example of a class of immunoglobulins.)
Physicochemical properties of proteins and methods for their separation.
6. How does the net charge of a protein affect its solubility?
a) determine the total charge of the peptide at pH 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis
b) how will the charge of this peptide change at pH >7, pH<7, рН <<7?
c) what is the isoelectric point of a protein (IEP) and in what environment does it lie?
IET of this peptide?
d) at what pH value will the lowest solubility of this peptide be observed.
7. Why does sour milk, unlike fresh milk, “curdle” when boiled (i.e., the milk protein casein precipitates)? In fresh milk, casein molecules have a negative charge.
8. Gel filtration is used to separate individual proteins. A mixture containing proteins A, B, C with molecular weights equal to 160,000, 80,000 and 60,000, respectively, was analyzed by gel filtration (Fig. 1.35). The swollen gel granules are permeable to proteins with a molecular weight of less than 70,000. What principle underlies this separation method? Which graph correctly reflects the results of fractionation? Indicate the order in which proteins A, B, and C are released from the column.
 Rice. 1.35. Using Gel Filtration to Separation of Proteins
Rice. 1.35. Using Gel Filtration to Separation of Proteins
9. In Fig. 1.36, A shows a diagram of electrophoresis on paper of blood serum proteins from a healthy person. The relative amounts of protein fractions obtained using this method are: albumins 54-58%, α 1 -globulins 6-7%, α 2 -globulins 8-9%, β-globulins 13%, γ-globulins 11-12% .
 Rice. 1.36 Electrophoresis on paper of blood plasma proteins of a healthy person (A) and a patient (B)
Rice. 1.36 Electrophoresis on paper of blood plasma proteins of a healthy person (A) and a patient (B)
I - γ-globulins; II - β-globulins; III -α 2 -globulin; IV -α 2 -globulin; V - albumins
Many diseases are accompanied by quantitative changes in the composition of serum proteins (dysproteinemia). The nature of these changes is taken into account when making a diagnosis and assessing the severity and stage of the disease.
Using the data given in table. 1.5, make a guess about the disease, which is characterized by the electrophoretic profile presented in Fig. 1.36.
Table 1.5. Changes in the concentration of serum proteins in pathology









- Experimental methods for recording elementary particles
- What is the Swedish match famous for?
- The best self-propelled howitzers Russian artillery
- Fedor Petrovich Litke: second circumnavigation of the world
- The first pharmacy was opened
- Wonderful traveler and scientist Fedor Petrovich Litke Stamp issued in honor of the Litke expedition
- Union of the Livonian Order with the Teutonic Order
- Pizza dough - secrets of Italian cuisine
- How to make shortcrust pastry baskets
- Layers of meat with lard in onion skins - it will work, you won’t be able to pull it off by the ears Boiled lard with a layer
- Pork gravy - recipes for pork gravy in a frying pan and in a slow cooker
- How to pay transport tax for legal entities
- Card for individual accounting of the amounts of accrued payments and other remunerations and the amounts of accrued insurance premiums Card for insurance premiums per year
- Error when filling in Contour
- Desk audit: developments
- Conditions for performing the stern experiment
- System status and processes
- Alexey Sergeevich Obukhov development of student research activities
- Mars 4th planet of the solar system
- Human organ systems