Ligand jest niezbędnym składnikiem złożonych białek. Pojęcie receptora i ligandu
Temat: TRANSMEMBRANA
Receptory wewnątrzkomórkowe: https://www.youtube.com/watch?v=Nm9u4lNCPyM
Metabotropowe receptory błonowe związane z systemem drugiego przekaźnika: https://www.youtube.com/watch?v=dQ4yVuLAbH0
Metabotropowe receptory błonowe związane z aktywnością kinazy tyrozynowej:
Rodzaje transportu przezbłonowego.
Rodzaje transbłonowych kanałów transportowych.
Pojęcie substancji polarnych i niepolarnych.
Selektywna przepuszczalność transbłonowa utrzymuje homeostazę komórkową, optymalną zawartość jonów, wody, enzymów i substratów w komórce. Sposoby realizacji selektywnej przepuszczalności błony: transport pasywny, transport katalizowany (dyfuzja ułatwiona), transport aktywny. Hydrofobowy charakter dwuwarstwowego rdzenia determinuje możliwość (lub niemożność) bezpośredniego przenikania przez membranę substancji różnych z fizykochemicznego punktu widzenia (głównie polarnych i niepolarnych).
Substancje niepolarne(na przykład cholesterol i jego pochodne) swobodnie przenikają przez błony biologiczne. Z tego powodu endocytoza i egzocytoza związków polarnych (na przykład hormonów peptydowych) zachodzi za pomocą pęcherzyków błonowych, a wydzielanie hormonów steroidowych zachodzi bez udziału takich pęcherzyków. Z tego samego powodu wewnątrz komórki znajdują się receptory cząsteczek niepolarnych (na przykład hormonów steroidowych).
Substancje polarne(np. białka i jony) nie mogą przenikać przez błony biologiczne. Dlatego wbudowane są receptory dla cząsteczek polarnych (na przykład hormonów peptydowych). błona plazmatyczna, a transmisja sygnału do innych przedziałów komórkowych odbywa się za pośrednictwem wtórnych przekaźników. Z tego samego powodu przezbłonowy transfer związków polarnych odbywa się za pomocą specjalnych systemów wbudowanych w błony biologiczne.
Selektywną przepuszczalność zapewnia błona komórkowa; Funkcję receptora realizują glikoproteiny, których części węglowodanowe znajdują się w glikokalisie, zachowanie kształtu i ruchliwość zapewniają białka fibrylarne i kanalikowe w warstwie podbłonowej itp.
Pojęcie receptora i ligandu
Receptor komórkowy- cząsteczka znajdująca się na powierzchni komórki, jądra, organelli komórkowych lub rozpuszczona w cytoplazmie. Receptor komórkowy reaguje specyficznie, zmieniając swoją konfigurację przestrzenną (kształt) na przyłączenie do niego cząsteczki określonej substancji chemicznej - ligand, przesyłając zewnętrzny sygnał regulacyjny. To z kolei przekazuje ten sygnał do komórki lub organelli komórkowych. Miejsce na receptorze, do którego się przyczepia ligand nazywany jest miejscem. Ten sam receptor może mieć kilka miejsc. Receptory komórkowe można podzielić na dwie główne klasy - receptory błonowe (znajduje się na błonie oddzielającej komórkę od środowiska zewnętrznego) i receptory wewnątrzkomórkowe.
Substancja, która specyficznie wiąże się z receptorem, nazywa się ligand (przez messengera) ten receptor. Zatem ligand (synonim: posłaniec) jest substancją chemiczną, która może oddziaływać z receptorem. Wynik tej interakcji może być różny. Jeśli ligand (posłaniec) prowadzi do zmiany kształtu receptora, a jego aktywacja nazywana jest agonistą . Jeśli ligand (posłaniec) zmieni kształt (konformację) receptora i blokują ten receptor, nazywa się to antagonistą.
Jeśli chodzi o zmysły, ligandy (posłańcy) to substancje działające na receptory węchu lub smaku.
Istnieją również termoczułe białka receptorowe i białka receptorowe, które reagują na zmiany potencjału błonowego.
Receptory dla rozpuszczalnych w wodzie ligandów (przekaźników) - hormonów białkowych, adrenaliny, noradrenaliny - znajdują się na powierzchni błony (receptory błonowe), wynika to z faktu, że ligandy hydrofilowe nie mogą przejść przez hydrofobową powierzchnię błony. Rozpuszczalne w tłuszczach ligandy (przekaźniki) z łatwością przechodzą przez dwuwarstwę fosfolipidową błony komórkowej i jądra, dlatego komórka lokalizuje dla nich receptory (receptory wewnątrzkomórkowe) wewnątrz: na organellach, w jądrze. Przykładami ligandów rozpuszczalnych w tłuszczach mogą być hormony steroidowe nadnerczy i gonad.
Ponadto ligandy można oddzielić do egzogennego(pochodzi z zewnątrz) i endogenny(powstaje wewnątrz ciała). Z reguły, jeśli jakaś egzogenna substancja ma receptory na komórkach, wówczas organizm ma również endogenne ligandy dla tego receptora. Przykładowo endogennym ligandem receptorów kannabinoidowych, z którymi wiążą się alkaloidy konopi indyjskich, jest substancja anandamid, wytwarzana przez organizm z arachidonowego kwasu tłuszczowego. Lub receptory endorfin (odgrywają ważną rolę w powstawaniu bólu i stan emocjonalny): mogą wiązać się z endogennymi ligandami - endorfinami lub mogą wiązać się z lekami z grupy morfin.
Wysoką specyficzność wiązania białka z ligandem zapewnia komplementarność struktury centrum aktywnego białka ze strukturą liganda
Komplementarność odnosi się do zgodności przestrzennej i chemicznej oddziałujących cząsteczek. Ligand musi mieć zdolność do wnikania i przestrzennego pokrywania się z konformacją miejsca aktywnego. Zbieżność ta może nie jest całkowita, ale ze względu na labilność konformacyjną białka, centrum aktywne jest zdolne do niewielkich zmian i jest „dopasowane” do liganda. Ponadto pomiędzy grupami funkcyjnymi liganda i rodnikami aminokwasowymi tworzącymi centrum aktywne muszą powstać wiązania utrzymujące ligand w centrum aktywnym. Wiązania pomiędzy ligandem a centrum aktywnym białka mogą być niekowalencyjne (jonowe, wodorowe, hydrofobowe) lub kowalencyjne.
1. Charakterystyka miejsca aktywnego
Centrum aktywne białka to region stosunkowo odizolowany od środowiska otaczającego białko, utworzony przez reszty aminokwasowe. W tym obszarze każda reszta, ze względu na swój indywidualny rozmiar i grupy funkcyjne, tworzy „relief” centrum aktywnego.
Połączenie takich aminokwasów w jeden kompleks funkcjonalny zmienia reaktywność ich rodników, tak jak zmienia się brzmienie instrumentu muzycznego w zespole. Dlatego reszty aminokwasowe tworzące centrum aktywne są często nazywane „zespołem” aminokwasów.
Unikalne właściwości centrum aktywnego zależą nie tylko od właściwości chemicznych tworzących go aminokwasów, ale także od ich dokładnej względnej orientacji w przestrzeni. Dlatego nawet niewielkie naruszenia ogólnej konformacji białka w wyniku punktowych zmian w jego pierwotnej strukturze lub warunkach środowisko może prowadzić do zmian właściwości chemicznych i funkcjonalnych rodników tworzących centrum aktywne, zakłócać wiązanie białka z ligandem i jego funkcję. Podczas denaturacji następuje zniszczenie centrum aktywnego białek i utrata ich aktywności biologicznej.
Często centrum aktywne jest utworzone w taki sposób, że dostęp wody do grup funkcyjnych jego rodników jest ograniczony, tj. powstają warunki do wiązania ligandu z rodnikami aminokwasów.
W niektórych przypadkach ligand przyłącza się tylko do jednego z atomów, który ma określoną reaktywność, na przykład dodanie O2 do żelaza mioglobiny lub hemoglobiny. Jednakże o właściwościach danego atomu do selektywnego oddziaływania z O2 decydują właściwości rodników otaczających atom żelaza w składzie. Hem występuje również w innych białkach, takich jak cytochromy. Jednak funkcja atomu żelaza w cytochromach jest inna: służy jako pośrednik w przenoszeniu elektronów z jednej substancji na drugą, podczas gdy żelazo staje się dwu- lub trójwartościowe.
Główną właściwością białek, która leży u podstaw ich funkcji, jest selektywność przyłączania określonych ligandów do określonych części cząsteczki białka.
2. Różnorodność ligandów
Ligandami mogą być substancje nieorganiczne (często jony metali) i organiczne, substancje o niskiej i wysokiej masie cząsteczkowej;
istnieją ligandy, które po przyłączeniu do miejsca aktywnego białka zmieniają swoją strukturę chemiczną (zmiany w substracie w miejscu aktywnym enzymu);
istnieją ligandy, które przyłączają się do białka dopiero w momencie funkcjonowania (na przykład O 2 transportowany przez hemoglobinę) oraz ligandy, które są stale związane z białkiem i odgrywają pomocniczą rolę w funkcjonowaniu białek (na przykład żelazo, który jest częścią hemoglobiny).
W przypadkach, gdy reszty aminokwasowe tworzące centrum aktywne nie mogą zapewnić funkcjonowania danego białka, cząsteczki niebiałkowe mogą przyłączać się do pewnych obszarów centrum aktywnego. Zatem centrum aktywne wielu enzymów zawiera jon metalu (kofaktor) lub organiczną cząsteczkę niebiałkową (koenzym). Część niebiałkowa, ściśle związana z centrum aktywnym białka i niezbędna do jego funkcjonowania, nazywana jest „grupą prostaty”. Mioglobina, hemoglobina i cytochromy mają grupę prostetyczną w centrum aktywnym - hem zawierający żelazo.
Połączenie protomerów w białku oligomerycznym jest przykładem oddziaływania ligandów o dużej masie cząsteczkowej. Każdy protomer, połączony z innymi protomerami, służy im jako ligand, tak samo jak oni dla niego.
Czasami przyłączenie ligandu zmienia konformację białka, powodując utworzenie miejsca wiązania z innymi ligandami. Na przykład białko kalmoduliny po związaniu się z czterema jonami Ca 2+ w określonych obszarach nabywa zdolność do interakcji z niektórymi enzymami, zmieniając ich aktywność.
8. Czwartorzędowa struktura białek. Cechy budowy i funkcjonowania białek oligomerycznych na przykładzie hemoglobiny. Spółdzielcze zmiany w konformacji protomeru. Możliwość regulacji funkcji biologicznej białek oligomerycznych przez ligandy allosteryczne.
Przez strukturę czwartorzędową rozumiemy metodę układania w przestrzeni pojedynczych łańcuchów polipeptydowych, które mają tę samą (lub inną) strukturę pierwszorzędową, drugorzędową lub trzeciorzędową i tworzenie strukturalnie i funkcjonalnie jednolitej formacji makromolekularnej. Wiele białek funkcjonalnych składa się z kilku łańcuchów polipeptydowych połączonych nie wiązaniami kowalencyjnymi, ale wiązaniami niekowalencyjnymi (podobnymi do tych, które zapewniają stabilność struktury trzeciorzędowej). Każdy pojedynczy łańcuch polipeptydowy, zwany protomerem, monomerem lub podjednostką, najczęściej nie wykazuje aktywności biologicznej. Białko nabywa tę zdolność poprzez pewien sposób przestrzennego asocjacji wchodzących w jego skład protomerów, tj. pojawia się nowa jakość, która nie jest charakterystyczna dla białka monomerycznego. Powstała cząsteczka jest zwykle nazywana oligomerem (lub multimerem). Białka oligomeryczne często zbudowane są z parzystej liczby protomerów (od 2 do 4, rzadziej od 6 do 8) o takich samych lub różnych masach cząsteczkowych – od kilku tysięcy do setek tysięcy. W szczególności cząsteczka hemoglobiny składa się z dwóch identycznych łańcuchów polipeptydowych α i dwóch β, tj. jest tetramerem.
Spółdzielcze zmiany w konformacji protomeru.
Zmiana konformacji, a co za tym idzie właściwości funkcjonalnych wszystkich protomerów białka oligomerycznego, gdy ligand jest przyłączony tylko do jednego z nich, nazywana jest kooperacyjnymi zmianami w konformacji protomerów.
Regulacja allosteryczna . Enzym moduluje swoją aktywność za pomocą niekowalencyjnie związanego z nim efektora. Wiązanie zachodzi w miejscu przestrzennie odległym od centrum aktywnego (katalitycznego). Wiązanie to powoduje zmiany konformacyjne w cząsteczce białka, co prowadzi do zmiany specyficznej geometrii centrum katalitycznego. Aktywność może wzrosnąć – jest to aktywacja enzymu, lub zmniejszyć – jest to hamowanie. „Wiadomość” o dodaniu aktywatora allosterycznego przekazywana jest poprzez zmiany konformacyjne do podjednostki katalitycznej, która staje się komplementarna do substratu, a enzym „ włącza się”. Po usunięciu aktywatora enzym ponownie przechodzi w formę nieaktywną i „wyłącza się”. Regulacja allosteryczna jest głównym sposobem regulacji szlaków metabolicznych.
Ppt%5C34928-slozhnye_belki_ch1_1.jpg" alt=">Centrum aktywne białka i jego oddziaływanie z ligandem. Podczas tworzenia struktury trzeciorzędowej"> Активный центр белка и его взаимодействие с лигандом. В процессе формирования третичной структуры на поверхности функционально активного белка, обычно в углублении, образуется участок, сформированный радикалами аминокислот, далеко стоящими друг от друга в первичной структуре. Этот участок, имеющий уникальное строение для данного белка и способный специфично взаимодействовать с определенной молекулой или группой похожих молекул, называется центром связывания белка с лигандом или активным центром. Лигандами называются молекулы, взаимодействующие с белками.!}
Ppt%5C34928-slozhnye_belki_ch1_2.jpg" alt=">Ligandem może być substancja o niskiej masie cząsteczkowej lub substancja o dużej masie cząsteczkowej (makrocząsteczka), w tym"> Лигандом может быть как низкомолекулярное, так и высокомолекулярное (макромолекула) вещество, в том числе и другой белок. Лигандами являются субстраты ферментов, кофакторы, ингибиторы и активаторы ферментов, протомеры в олигомерном белке и т.д.!}
Ppt%5C34928-slozhnye_belki_ch1_3.jpg" alt=">Wysoką specyficzność interakcji białko-ligand zapewnia komplementarność struktury centrum aktywnego ze strukturą liganda.">!}
Ppt%5C34928-slozhnye_belki_ch1_4.jpg" alt=">Komplementarność to przestrzenna i chemiczna zgodność oddziałujących powierzchni. Aktywny ośrodek musi nie tylko"> Комплементарность - это пространственное и химическое соответствие взаимодействующих поверхностей. Активный центр должен не только пространственно соответствовать входящему в него лиганду, но и между функциональными группами радикалов, входящих в активный центр, и лигандом должны образоваться связи чаще всего нековалентные (ионные, водородные, а также гидрофобные взаимодействия), которые удерживают лиганд в активном центре.!}
Ppt%5C34928-slozhnye_belki_ch1_5.jpg" alt=">Uzupełniające oddziaływanie białka z ligandem">!}
Ppt%5C34928-slozhnye_belki_ch1_6.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_7.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_8.jpg" alt="> KLASYFIKACJA BIAŁEK 1. Białka proste składają się wyłącznie z aminokwasów. 2. Białka złożone (holoproteiny)"> КЛАССИФИКАЦИЯ БЕЛКОВ 1. Простые белки состоят только из аминокислот. 2. Сложные белки (холопротеины) содержат белковую часть (апопротеин) и небелковую (простетическую) группу.!}
Ppt%5C34928-slozhnye_belki_ch1_9.jpg" alt=">Różne substancje organiczne (lipidy, węglowodany) i nieorganiczne (metale) mogą działać jako grupa prostetyczna."> В качестве простетической группы могут выступать различные органические (липиды, углеводы) и неорганические (металлы) вещества. Связь между простетической группой и апопротеином может быть как ковалентная, так и нековалентная. Простетическую группу порой можно рассматривать в качестве лиганда. Наличие небелковой части обеспечивает выполнение белком его функции. При утрате простетической группы холопротеин теряет свою активность.!}
Ppt%5C34928-slozhnye_belki_ch1_10.jpg" alt=">Białka złożone - chromoproteiny - nukleoproteiny - lipoproteiny - fosfoproteiny - glikoproteiny - metaloproteiny">!}
Ppt%5C34928-slozhnye_belki_ch1_11.jpg" alt=">Do metaloprotein zalicza się holoenzymy zawierające jony metali niehemowe skoordynowane. Do metaloprotein zaliczają się białka"> Металлопротеинам можно отнести холоферменты, содержащие негемовые координационно связанные ионы металлов. Среди металлопротеинов есть белки, выполняющие депонирующие и транспортные функции (например, железосодержащие ферритин и трансферрин) и ферменты (например, цинксодержащая карбоангидраза и различные супероксиддисмутазы, содержащие в качестве активных центров ионы меди, марганца, железа и других металлов). Но и хромопротеины, содержащие ионы металлов, также можно отнести к металлопротеинам.!}
Ppt%5C34928-slozhnye_belki_ch1_12.jpg" alt=">Metaloproteiny są często enzymami. Jony metali w tym przypadku: - uczestniczą w orientacji podłoża"> Металлопротеины часто являются ферментами. Ионы металлов в этом случае: - участвуют в ориентации субстрата в активном центре фермента, входят в состав активного центра фермента и участвуют в катализе, являясь, например, акцепторами электронов на определенной стадии ферментативной реакции. Часто ион металла в составе фермента называют кофактором.!}
Ppt%5C34928-slozhnye_belki_ch1_13.jpg" alt=">Do metaloprotein enzymatycznych zalicza się białka zawierające w kompleksie np.: - miedź - oksydazę cytochromową"> К ферментативным металлопротеинам относятся белки, содержащие например: - медь – цитохромоксидаза, в комплексе с другими ферментами дыхательной цепи митохондрий участвует в синтезе АТФ, - железо – ферритин, депонирующий железо в клетке, трансферрин, переносящий железо в крови, каталаза, обезвреживающая перекись водорода, - цинк – алкогольдегидрогеназа, обеспечивающая метаболизм этанола и других спиртов, лактатдегидрогеназа, участвующая в метаболизме молочной кислоты, - карбоангидраза, образующая угольную кислоту из CO2 и H2O, - щелочная фосфатаза, гидролизующая фосфорные эфиры различных соединений, - α2-макроглобулин, антипротеазный белок крови. - селен – тиреопероксидаза, участвующая в синтезе гормонов !} Tarczyca, enzym przeciwutleniający peroksydaza glutationowa, - wapń - α-amylaza śliny i soku trzustkowego, hydrolizująca skrobię.
Ppt%5C34928-slozhnye_belki_ch1_14.jpg" alt=">Ferrytyna">!}
Ppt%5C34928-slozhnye_belki_ch1_15.jpg" alt=">Fosfoproteiny to białka zawierające grupę fosforanową. Wiąże się ona z łańcuchem peptydowym"> Фосфопротеины – это белки, в которых присутствует фосфатная группа. Она связывается с пептидной цепью через остатки тирозина, серина и треонина, т.е. тех аминокислот, которые содержат ОН-группу. Способ присоединения фосфата к белку на примере серина и тирозина!}
Ppt%5C34928-slozhnye_belki_ch1_16.jpg" alt=">Kwas fosforowy może pełnić: - rolę strukturalną, nadając ładunek, rozpuszczalność i zmieniając właściwości"> Фосфорная кислота может выполнять: - Структурную роль, придавая заряд, растворимость и изменяя свойства белка, например, в казеине молока, яичном альбумине. Наличие остатков фосфорной кислоты способствует связыванию кальция, что необходимо для формирования, например, костной ткани. - Функциональную роль. В клетке присутствует много белков, которые связаны с фосфатом не постоянно, а в зависимости от активности метаболизма. Белок может многократно переходить в фосфорилированную или в дефосфорилированную форму, что играет регулирующую роль в его работе.!}
Ppt%5C34928-slozhnye_belki_ch1_17.jpg" alt=">Fosforylacja to proces przeniesienia reszty kwasu fosforowego z fosforylującego donora na substrat, zwykle"> Фосфорилирование - процесс переноса остатка фосфорной кислоты от фосфорилирующего агента-донора к субстрату, как правило, катализируемый ферментами (киназами) и ведущий к образованию эфиров фосфорной кислоты. Дефосфорилирование (утрату остатка фосфорной кислоты) катализируют фосфатазы. АТФ + R-OH → АДФ + R-OPO3H2 R-OPO3H2 + Н2О → R-OH + Н3РО4!}
Ppt%5C34928-slozhnye_belki_ch1_18.jpg" alt=">Przykłady: 1) enzymy syntaza glikogenu i fosforylaza glikogenu 2) histony w stanie fosforylowanym wiążą się słabiej"> Примеры: 1) ферменты гликогенсинтаза и гликогенфосфорилаза 2) гистоны в фосфорилированном состоянии менее прочно связываются с ДНК и активность генома возрастает. Изменение конформации белка в фосфорилированном и дефосфорилированном состоянии!}
Ppt%5C34928-slozhnye_belki_ch1_19.jpg" alt=">Lipoproteiny zawierają niekowalencyjnie związane lipidy jako część protetyczną. W szczególności lipidy"> Липопротеины содержат в качестве простетической части нековалентно связанные липиды. Липиды, в частности жиры, холестерол и его эфиры не растворяются в водных фазах организма, поэтому транспорт их кровью и лимфой осуществляется в виде комплексов с белками и фосфолипидами, которые называются липопротеинами.!}
Ppt%5C34928-slozhnye_belki_ch1_20.jpg" alt=">Wszystkie lipoproteiny mają podobną budowę: rdzeń składa się z cząsteczek hydrofobowych: triacylogliceroli, estrów cholesterolu i"> Все липопротеины имеют сходное строение: ядро состоит из гидрофобных молекул: триацилглицеролов, эфиров холестерола, а на поверхности находится монослой фосфолипидов, полярные группы которых обращены к воде, а гидрофобные погружены в гидрофобное ядро липопротеина. Кроме фосфолипидов, на поверхности находятся белки – аполипопротеины (апобелками). Их выделяют несколько видов: А, В, С, D. В каждом типе липопротеинов преобладают соответствующие ему апобелки. Аполипопротеины выполняют различные функции. Интегральные аполипопротеины являются структурными компонентами. Периферические аполипопротеины в плазме крови могут передаваться от одного типа липопротеинов к другим, определяя их дальнейшие превращения.!}
Ppt%5C34928-slozhnye_belki_ch1_21.jpg" alt=">Schemat struktury lipoproteiny Struktura lipoproteiny">!}
Ppt%5C34928-slozhnye_belki_ch1_22.jpg" alt=">Struktura lipoprotein w osoczu krwi">!}
Ppt%5C34928-slozhnye_belki_ch1_23.jpg" alt=">Istnieją cztery główne klasy lipoprotein: - lipoproteiny o dużej gęstości (HDL), - lipoproteiny o małej gęstości (LDL),"> Выделяют четыре основных класса липопротеинов: -липопротеины высокой плотности (ЛПВП), -липопротеины низкой плотности (ЛПНП), -липопротеины очень низкой плотности (ЛПОНП), -хиломикроны (ХМ). Каждый из типов ЛП образуется в разных тканях и транспортирует определённые липиды. Концентрация и соотношение в крови тех или иных липопротеинов играют ведущую роль в возникновении такой распространенной сосудистой патологии как атеросклероз. ЛПВП являются антиатерогенными, ЛПНП и ЛПОНП – атерогенными.!}
Ppt%5C34928-slozhnye_belki_ch1_24.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_25.jpg" alt=">Glikoproteiny lub glikokoniugaty to białka zawierające składnik węglowodanowy kowalencyjnie przyłączony do szkieletu polipeptydowego."> Гликопротеины или, гликоконъюгаты – это белки, содержащие углеводный компонент, ковалентно присоединенный к полипептидной основе. Содержание углеводов в них варьирует от 1% до 98% по массе. Два подкласса белков, содержащих углеводы: ■ протеогликаны ■ гликопротеины!}
Opis="">
Ppt%5C34928-slozhnye_belki_ch1_27.jpg" alt=">Glikoproteiny charakteryzują się niską zawartością węglowodanów, które są przyłączone: - wiązaniem N-glikozydowym z grupą NH2 niektórych"> Для гликопротеинов характерно невысокое содержание углеводов, которые присоединены: - N-гликозидной связью к NН2-группе какого-нибудь аминокислотного остатка, например, аспарагина; - О-гликозидной связью к гидроксильной группе остатка серина, треонина,тирозина, гидроксилизина.!}
Ppt%5C34928-slozhnye_belki_ch1_28.jpg" alt=">Tworzenie wiązań O- i N-glikozydowych w glikoproteinach. 1 - Wiązanie N-glikozydowe pomiędzy grupą amidową"> Образование О- и N-гликозидных связей в гликопротеинах. 1 - N-гликозидная связь между амидной группой аспарагина и ОН-группой моносахарида; 2 - О-гликозидная связь между ОН-группой серина и ОН-группой моносахарида.!}
Ppt%5C34928-slozhnye_belki_ch1_29.jpg" alt=">Sposób dodawania węglowodanów do białka">!}
Ppt%5C34928-slozhnye_belki_ch1_30.jpg" alt=">Funkcje glikoprotein to: 1. Strukturalne - ściana komórkowa bakterii, macierz kostna, np. kolagen, elastyna."> Функцией гликопротеинов являются: 1. Структурная – клеточная стенка бактерий, костный матрикс, например, коллаген, эластин. 2. Защитная – например, антитела, интерферон, факторы свертывания крови (протромбин, фибриноген). 3. Рецепторная – присоединение эффектора приводит к изменению конформации белка-рецептора, что вызывает внутриклеточный ответ. 4. Гормональная – гонадотропный, адренокортикотропный и тиреотропный гормоны. 5. Ферментативная – холинэстераза, нуклеаза. 6. Транспортная – перенос веществ в крови и через мембраны, например, трансферрин, транскортин, альбумин, Na+,К+-АТФаза.!}
Ppt%5C34928-slozhnye_belki_ch1_31.jpg" alt=">Schemat struktury białka receptorowego">!}
Ppt%5C34928-slozhnye_belki_ch1_32.jpg" alt=">Chromoproteiny - nazwa zbiorcza złożone białka z kolorowymi grupami protetycznymi o różnym charakterze chemicznym. "> Chromoproteiny to zbiorcza nazwa złożonych białek z kolorowymi grupami prostetycznymi o różnym charakterze chemicznym. hemoproteiny (zawierają hem), białka siatkówki (zawierają witaminę A), flawoproteiny (zawierają witaminę B2), białka kobamidu (zawierają witaminę B12).
Ppt%5C34928-slozhnye_belki_ch1_33.jpg" alt=">Flawoproteiny to enzymy reakcji redoks. Zawierają pochodne witaminy B2, mononukleotyd flawinowy (FMN) i dinukleotyd flawinowo-adeninowy"> Флавопротеины - это ферменты окислительно-восстановительных реакций. Содержат производные витамина В2 флавинмононуклеотид (ФМН) и флавинадениндинуклеотид (ФАД). Связываются данные простетические группы ковалентно и придают желтое окрашивание. Эти простетические группы являются производными изоаллоксазина.!}
Ppt%5C34928-slozhnye_belki_ch1_34.jpg" alt=">Izoalloksazyna jest związkiem heterocyklicznym, pochodną pterydyny. Cząsteczka izoalloksazyny składa się z trzech pierścieni aromatycznych -"> Изоаллоксазин - гетероциклическое соединения, производное птеридина. Молекула изоаллоксазина состоит из трех ароматических колец - бензольного, пиримидинового, пиразинового.!}
Ppt%5C34928-slozhnye_belki_ch1_35.jpg" alt=">Hemoproteiny to chromoproteiny zawierające hem. Zawierają one strukturalnie podobne porfiryny żelaza lub magnezu jako składnik niebiałkowy."> Гемопротеины - гем-содержащие хромопротеины. В качестве небелкового компонента включают структурно сходные железо- или магнийпорфирины. Белковый компонент может быть разнообразным как по составу, так и по структуре. Основу структуры простетической группы большинства гемосодержащих белков составляет порфириновое кольцо, являющееся в свою очередь производным тетрапиррольного соединения – порфирина. Порфирин!}
Ppt%5C34928-slozhnye_belki_ch1_36.jpg" alt=">Pierścień porfirynowy jest zdolny do tworzenia związków koordynacyjnych z jonami różnych metali. W wyniku kompleksowania,"> Порфириновое кольцо способно образовывать координационные соединения с различными ионами металлов. В результате комплексообразования формируются металлопорфирины: содержащие ионы железа – гемоглобины, миоглобин, цитохромы, пероксидаза, каталаза и др. (красное окрашивание), содержщие ионы магния – хлорофилл (зеленое окрашивание). Витамин В12 (кобалимин) содержит координированный ион кобальта Со2+ в порфириноподобном макроцикле – коррине, состоящем из четырех частично гидрированных пиррольных колец (розовое окрашивание).!}
Ppt%5C34928-slozhnye_belki_ch1_37.jpg" alt=">Chlorofil b. Chlorofile biorą udział w procesach fotosyntezy.">!}
Ppt%5C34928-slozhnye_belki_ch1_38.jpg" alt=">Cytochromy różnią się składem aminokwasowym łańcuchów peptydowych, liczbą łańcuchów i dzielą się na typy a, b,"> Цитохромы различаются аминокислотным составом пептидных цепей, числом цепей и разделяются на типы а, b, с, d. Цитохромы находятся в составе дыхательной цепи и цепи микросомального окисления. Степень окисления железа в составе цитохромов меняется в отличие от гемоглобина и миоглобина Fe2+ ↔ Fe3+!}
Ppt%5C34928-slozhnye_belki_ch1_39.jpg" alt=">Mioglobina (MB) to białko występujące w czerwonych mięśniach, którego główną funkcją jest tworzenie rezerw"> Миоглобин (Мв) - белок, находящийся в красных мышцах, основная функция которого - создание запасов О2, необходимых при интенсивной мышечной работе. Мв - сложный белок, содержащий белковую часть - апоМв и небелковую часть - гем. Первичная структура апоМв определяет его компактную глобулярную конформацию и структуру активного центра, к которому присоединяется небелковая часть миоглобина - гем. Кислород, поступающий из крови в мышцы, связывается с Fe2+ гема в составе миоглобина. Мв - мономерный белок, имеющий очень высокое сродство к О2, поэтому отдача кислорода миоглобином происходит только при интенсивной мышечной работе, когда парциальное давление O2 резко снижается. Формирование пространственных структур и функционирование миоглобина.!}
Ppt%5C34928-slozhnye_belki_ch1_40.jpg" alt=">Tworzenie konformacji MV. W mięśniach czerwonych synteza białka pierwotnego zachodzi na rybosomach podczas translacji"> Формирование конформации Мв. В красных мышцах на рибосомах в ходе трансляции идет синтез первичной структуры Мв, представленной специфической последовательностью 153 аминокислотных остатков. Вторичная структура Мв содержит восемь α-спиралей, называемых латинскими буквами от А до Н, между которыми имеются неспирализованные участки. Третичная структура Мв имеет вид компактной глобулы, в углублении которой между F и Е α-спиралями расположен активный центр.!}
Ppt%5C34928-slozhnye_belki_ch1_41.jpg" alt=">Struktura mioglobiny">!}
Ppt%5C34928-slozhnye_belki_ch1_42.jpg" alt=">Cechy budowy i funkcjonowania centrum aktywnego Mv. Centrum aktywne Mv zbudowane jest głównie z rodników hydrofobowych"> Особенности строения и функционирования активного центра Мв. Активный центр Мв сформирован преимущественно гидрофобными радикалами аминокислот, далеко отстоящими друг от друга в первичной структуре (например, Три39 и Фен138). К активному центру присоединяется плохо растворимые в воде лиганды - гем и О2. Гем - специфический лиганд апоМв.!}
Ppt%5C34928-slozhnye_belki_ch1_43.jpg" alt=">Podstawę hemu tworzą cztery pierścienie pirolowe połączone mostkami metylowymi; w środku znajduje się atom Fe2+,"> Основу гема составляют четыре пиррольных кольца, соединенных метенильными мостиками; в центре расположен атом Fe2+, соединенный с атомами азота пиррольных колец четырьмя координационными связями. В активном центре Мв кроме гидрофобных радикалов аминокислот имеются также остатки двух аминокислот с гидрофильными радикалами - Гис Е7 (Гис64) и Гис F8 (Гис93).!}
Ppt%5C34928-slozhnye_belki_ch1_44.jpg" alt=">Jego F8 tworzy wiązanie koordynacyjne z Fe2+ i mocno wiąże hem w miejscu aktywnym."> Гис F8 образует координационную связь с Fe2+ и прочно фиксирует гем в активном центре. Гис Е7 необходим для правильной ориентации в активном центре другого лиганда - O2 при его взаимодействии с Fe+2 гема. Микроокружение гема создает условия для прочного, но обратимого связывания O2 с Fe+2 и препятствует попаданию в гидрофобный активный центр воды, что может привести к его окислению в Fе3+.!}
Ppt%5C34928-slozhnye_belki_ch1_45.jpg" alt=">Struktura oligomeryczna HB i regulacja powinowactwa HB do O2 przez ligandy. Hemoglobiny ludzkie -"> Олигомерное строение Нв и регуляция сродства Нв к О2 лигандами. Гемоглобины человека - семейство белков, так же как и миоглобин относящиеся к сложным белкам (гемопротеинам). Они имеют тетрамерное строение и содержат две α-цепи, но различаются по строению двух других полипептидных цепей (2α-, 2х-цепи). Строение второй полипептидной цепи определяет особенности функционирования этих форм Нв. Около 98% гемоглобина эритроцитов взрослого человека составляет гемоглобин А (2α-, 2β-цепи). В период внутриутробного развития функционируют два основных типа гемоглобинов: эмбриональный Нв (2α, 2ε), который обнаруживается на ранних этапах развития плода, и гемоглобин F (фетальный) - (2α, 2γ), который приходит на смену раннему гемоглобину плода на шестом месяце внутриутробного развития и только после рождения замещается на Нв А.!}
Ppt%5C34928-slozhnye_belki_ch1_46.jpg" alt=">Hb A to białko spokrewnione z mioglobiną (Mb), występujące w czerwonych krwinkach osoby dorosłej. Jego struktura"> Нв А - белок, родственный миоглобину (Мв), содержится в эритроцитах взрослого человека. Строение его отдельных протомеров аналогично таковому у миоглобина. Вторичная и третичная структуры миоглобина и протомеров гемоглобина очень сходны, несмотря на то что в первичной структуре их полипептидных цепей идентичны только 24 аминокислотных остатка (вторичная структура протомеров гемоглобина, так же как миоглобин, содержит восемь α-спиралей, обозначаемых латинскими буквами от А до Н, а третичная структура имеет вид компактной глобулы). Но в отличие от миоглобина гемоглобин имеет олигомерное строение, состоит из четырех полипептидных цепей, соединенных нековалентными связями.!}
Ppt%5C34928-slozhnye_belki_ch1_47.jpg" alt=">Oligomeryczna struktura hemoglobiny">!}
Ppt%5C34928-slozhnye_belki_ch1_48.jpg" alt=">Każdy protomer Hb jest powiązany z częścią niebiałkową - hemem i sąsiednimi protomerami. Połączenie białka"> Каждый протомер Нв связан с небелковой частью - гемом и соседними протомерами. Соединение белковой части Нв с гемом аналогично таковому у миоглобина: в активном центре белка гидрофобные части гема окружены гидрофобными радикалами аминокислот за исключением Гис F8 и Гис Е7, которые расположены по обе стороны от плоскости гема и играют аналогичную роль в функционировании белка и связывании его с кислородом. Кроме того, Гис Е7 выполняет важную дополнительную роль в функционировании Нв. Свободный гем имеет в 25 000 раз более высокое сродство к СО, чем к О2. СО в небольших количествах образуется в организме и, учитывая его высокое сродство к гему, он мог бы нарушать транспорт необходимого для жизни клеток О2. Однако в составе гемоглобина сродство гема к оксиду углерода превышает сродство к О2 всего в 200 раз благодаря наличию в активном центре Гис Е7. Остаток этой аминокислоты создает оптимальные условия для связывания гема с O2 и ослабляет взаимодействие гема с СО.!}
Ppt%5C34928-slozhnye_belki_ch1_49.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_50.jpg" alt=">Pierścienie pirolowe hemu znajdują się w tej samej płaszczyźnie, a jon Fe2+ jest w stanie nieutlenionym Hb"> Пиррольные кольца гема расположены в одной плоскости, а ион Fe2+ в неоксигенированом состоянии Hb выступает над плоскостью на 0,6 А. При присоединении кислорода ион железа погружается в плоскость колец гема. В результате сдвигается и участок полипептидной цепи, нарушаются слабые связи в молекуле Hb и изменяется конформация всей глобулы. Таким образом, присоединение кислорода вызывает изменение пространственной структуры молекулы миоглобина или протомеров гемоглобина.!}
Ppt%5C34928-slozhnye_belki_ch1_51.jpg" alt=">Hemoglobina może występować zarówno w postaci wolnej (deoksyhemoglobina), jak i utlenionej, sumując się do"> Гемоглобин может существовать как в свободной (дезоксигемоглобин), так и в оксигенированной форме, присоединяя до 4 молекул кислорода. Взаимодействие с кислородом 1-го протомера вызывает изменение его конформации, а также кооперативные конформационные изменения остальных протомеров. Сродство к кислороду возрастает, и присоединение О2 к активному центру 2-го протомера происходит легче, вызывая дальнейшую конформационную перестройку всей молекулы. В результате еще сильнее изменяется структура оставшихся протомеров и их активных центров, взаимодействие с О2 еще больше облегчается. В итоге 4-я молекула кислорода присоединяется к Hb примерно в 300 раз легче, чем 1-я. Так происходит в легких при высоком парциальном давлении кислорода.!}
Ppt%5C34928-slozhnye_belki_ch1_52.jpg" alt=">Wspólne zmiany w konformacji cząsteczki hemoglobiny pod wpływem interakcji z tlenem">!}
Ppt%5C34928-slozhnye_belki_ch1_53.jpg" alt=">W tkankach, w których zawartość tlenu jest mniejsza, wręcz przeciwnie, rozszczepienie każdej cząsteczki O2 ułatwia uwolnienie kolejnych."> В тканях, где содержание кислорода ниже, наоборот, отщепление каждой молекулы О2 облегчает освобождение последующих. Таким образом, взаимодействие олигомерного белка гемоглобина с лигандом (О2) в одном центре связывания приводит к изменению конформации всей молекулы и других, пространственно удаленных центров, расположенных на других субъединицах (принцип «домино»). Подобные взаимосвязанные изменения структуры белка называют кооперативными конформационными изменениями. Они характерны для всех олигомерных белков и используются для регуляции их активности.!}
Ppt%5C34928-slozhnye_belki_ch1_54.jpg" alt=">Oddziaływanie obu białek (Mb i Hb) z tlenem zależy od jego ciśnienia parcjalnego w"> Взаимодействие обоих белков (Mb и Hb) с кислородом зависит от его парциального давления в тканях. Эта зависимость имеет разный характер, что связано с их особенностями структуры и функционирования. Гемоглобин имеет S-образную кривую насыщения, которая показывает, что субъединицы белка работают кооперативно, и чем больше кислорода они отдают, тем легче идет освобождение остальных молекул О2. Этот процесс зависит от изменения парциального давления кислорода в тканях. График насыщения миоглобина кислородом имеет характер простой гиперболы, т.е. насыщение Mb кислородом происходит быстро и отражает его функцию - обратимое связывание с кислородом, высвобождаемым гемоглобином, и освобождение в случае интенсивной физической нагрузки.!}
Ppt%5C34928-slozhnye_belki_ch1_55.jpg" alt=">Krzywe nasycenia mioglobiny i hemoglobiny tlenem">!}
Ppt%5C34928-slozhnye_belki_ch1_56.jpg" alt=">CO2 i H+ powstające podczas katabolizmu substancji organicznych, zmniejszają proporcjonalnie powinowactwo hemoglobiny do O2"> CO2 и Н+, образующиеся при катаболизме органических веществ, уменьшают сродство гемоглобина к О2 пропорционально их концентрации. Энергия, необходимая для работы клеток, вырабатывается преимущественно в митохондриях при окислении органических веществ с использованием O2, доставляемого из легких гемоглобином. В результате окисления органических веществ образуются конечные продукты их распада: СО2 и Н2O, количество которых пропорционально интенсивности протекающих процессов окисления. СO2 диффузией попадает из клеток в кровь и проникает в эритроциты, где под действием фермента карбоангидразы превращается в угольную кислоту. Эта слабая кислота диссоциирует на протон и бикарбонат ион. СО2 + Н2О → Н2СО3 → Н+ + НСО3-!}
Ppt%5C34928-slozhnye_belki_ch1_57.jpg" alt=">Jony H+ potrafią łączyć się z rodnikami His146 w łańcuchach β hemoglobiny, tj. w obszarach odległych"> Ионы Н+ способны присоединятся к радикалам Гис146 в β-цепях гемоглобина, т.е. в участках, удаленных от гема. Протонирование гемоглобина снижает его сродство к О2, способствует отщеплению О2 от оксиНв, образованию дезоксиНв и увеличивает поступление кислорода в ткани пропорционально количеству образовавшихся протонов. Увеличение количества освобожденного кислорода в зависимости от увеличения концентрации Н+ в эритроцитах называется эффектом Бора (по имени датского физиолога Христиана Бора, впервые открывшего этот эффект). В легких высокое парциальное давление кислорода способствует его связыванию с дезоксиНв, что уменьшает сродство белка к Н+. Освободившиеся протоны под действием карбоангидразы взаимодействуют с бикарбонатами с образованием СО2 и Н2О!}
Ppt%5C34928-slozhnye_belki_ch1_58.jpg" alt=">Zależność powinowactwa Hb do O2 od stężenia CO2 i protonów (efekt Bohra): A -"> Зависимость сродства Нв к О2 от концентрации СО2 и протонов (эффект Бора): А - влияние концентрации СО2 и Н+ на высвобождение О2 из комплекса с Нв (эффект Бора); Б - оксигенирование дезоксигемоглобина в легких, образование и выделение СО2.!}
Ppt%5C34928-slozhnye_belki_ch1_59.jpg" alt=">Powstający CO2 przedostaje się do przestrzeni pęcherzykowej i jest usuwany z wydychanym powietrzem. Zatem ilość"> Образовавшийся СО2 поступает в альвеолярное пространство и удаляется с выдыхаемым воздухом. Таким образом, количество высвобождаемого гемоглобином кислорода в тканях регулируется продуктами катаболизма органических веществ: чем интенсивнее распад веществ, например при физических нагрузках, тем выше концентрация СО2 и Н+ и тем больше кислорода получают ткани в результате уменьшения сродства Нв к О2.!}
Ppt%5C34928-slozhnye_belki_ch1_60.jpg" alt=">Zmiana aktywności funkcjonalnej białka podczas interakcji z innymi ligandami w wyniku zmian konformacyjnych nazywana jest allosteryczną"> Изменение функциональной активности белка при взаимодействии с другими лигандами вследствие конформационных изменений называется аллостерической регуляцией, а соединения-регуляторы - аллостерическими лигандами или эффекторами. Способность к аллостерической регуляции характерна, как правило, для олигомерных белков, т.е. для проявления аллостерического эффекта необходимо взаимодействие протомеров. При воздействии аллостерических лигандов белки меняют свою конформацию (в том числе и активного центра) и функцию.!}
Ppt%5C34928-slozhnye_belki_ch1_61.jpg" alt=">Allosteryczna regulacja powinowactwa Hb do O2 przez ligand - 2,3-bis-fosfoglicerynian. W erytrocytach z produktu"> Аллостерическая регуляция сродства Нв к О2 лигандом - 2,3-бис-фосфоглицератом. В эритроцитах из продукта окисления глюкозы - 1,3-бисфосфоглицерата синтезируется аллостерический лиганд гемоглобина - 2,3-бисфосфоглицерат (2,3-БФГ). В нормальных условиях концентрация 2,3-БФГ высокая и сравнима с концентрацией Нв. 2,3-БФГ имеет сильный отрицательный заряд (-5).!}
Ppt%5C34928-slozhnye_belki_ch1_62.jpg" alt=">W środku tetramerycznej cząsteczki hemoglobiny znajduje się wnęka. Jest ona utworzona przez reszty aminokwasowe wszystkich czterech protomerów."> В центре тетрамерной молекулы гемоглобина находится полость. Ее образуют аминокислотные остатки всех четырех протомеров. В капиллярах тканей протонирование Нв (эффект Бора) приводит к разрыву связи между железом гема и О2. В молекуле дезоксигемоглобина по сравнению с оксигемоглобином возникают дополнительные ионные связи, соединяющие протомеры, вследствие чего размеры центральной полости по сравнению с оксигемоглобином увеличиваются. Центральная полость является местом присоединения 2,3-БФГ к гемоглобину. БФГ поступает в полость дезоксигемоглобина. 2,3-БФГ взаимодействует с гемоглобином в участке, удаленном от активных центров белка и относится к аллостерическим (регуляторным) лигандам, а центральная полость Нв является аллостерическим центром. 2,3-БФГ имеет сильный отрицательный заряд и взаимодействует с положительно заряженными группами двух β-цепей Нв. При этом его сродство к О2 снижается в 26 раз. В результате происходит высвобождение кислорода в капиллярах ткани при низком парциальном давлении О2. В легких высокое парциальное давление О2, наоборот, приводит к оксигенированию Нв и освобождению БФГ.!}
Ppt%5C34928-slozhnye_belki_ch1_63.jpg" alt=">Miejsce wiązania BPG znajduje się w dodatnio naładowanej wnęce pomiędzy 4 protomerami hemoglobiny. Interakcja BPG"> Центр связывания БФГ находится в положительно заряженной полости между 4 протомерами гемоглобина. Взаимодействие БФГ с центром связывания изменяет конформацию α- и β-протомеров НЬ и их активных центров. Сродство НЬ к молекулам О2 снижается и кислород высвобождается в ткани. В легких при высоком парциальном давлении О2 активные центры гемоглобина насыщаются за счет изменения конформации и БФГ вытесняется из аллостерического центра!}
Ppt%5C34928-slozhnye_belki_ch1_64.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_65.jpg" alt=">Dzięki temu białka oligomeryczne mają nowe właściwości w porównaniu z białkami monomerycznymi. Przyłączenie ligandów"> Таким образом, олигомерные белки обладают новыми по сравнению с мономерными белками свойствами. Присоединение лигандов на участках, пространственно удаленных друг от друга (аллостерических), способно вызывать конформационные изменения во всей белковой молекуле. Благодаря взаимодействию с регуляторными лигандами происходит изменение конформации и адаптация функции белковой молекулы к изменениям окружающей среды.!}
Ppt%5C34928-slozhnye_belki_ch1_66.jpg" alt=">Około 15% dwutlenku węgla obecnego we krwi jest przenoszone przez cząsteczki hemoglobiny. W tkankach część cząsteczek"> Около 15% углекислого газа, присутствующего в крови, переносится молекулами гемоглобина. В тканях часть молекул углекислого газа может присоединится к каждому протомеру молекулы гемоглобина, при этом снижается сродство Hb к кислороду. В легких, наоборот, из-за высокого парциального давления кислорода, О2 связывается с Hb, а СО2 высвобождается.!}
Ppt%5C34928-slozhnye_belki_ch1_67.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_68.jpg" alt=">W cząsteczce hemoglobiny S (tzw. hemoglobiny nieprawidłowej) zmutowane okazały się 2 łańcuchy β, w których"> В молекуле гемоглобина S (так назван аномальный гемоглобин) мутантными оказались 2 β-цепи, в которых глутамат, высокополярная отрицательно заряженная аминокислота в положении 6 была заменена валином, содержащим гидрофобный радикал.!}
Lub rodniki związane z centrum. atom złożonego związku. Mogą. jony (H - , Hal - , NO 3 - , NCS - itp.), inorg. cząsteczki (H 2, C n, N 2, P n, O 2, Sn, CO, CO 2, NH 3, NO, SO 2, NO 2, COS itp.), org. związki zawierające pierwiastki z głównych podgrup V, VI, VII gr. okresowy systemy lub funkcja p-donora. Duża grupa L.-biologicznie ważnych związków. (peptydy, puryn, korryn) i ich syntetyki. analogi (etery koronowe, ), a także z atomami donorowymi i grupami chelatującymi. L. można podłączyć do środka. atom z wiązaniami dwu- lub wielocentrowymi s-, p- i d-. W przypadku tworzenia się wiązań dwucentrowych w aluminium, można zidentyfikować centra donorowe (zwykle N, O, S, Cl lub atomy). Wiązanie wieloośrodkowe odbywa się dzięki aromatycznemu systemowi p. L. (anion cyklopentadienidowy) lub heteroaromatyczny. L. (tiofen, metylopirydyny). Najważniejsza ilość. cechą charakterystyczną zdolności więzadła dawca-akceptor jest uzębienie, określone przez liczbę ośrodków dawcy więzadła zaangażowanych w koordynację. Na tej podstawie L. dzieli się na mono-, di-, ... polidentat. Koordynat. liczba czynnika kompleksującego dla więzadeł jednokleszczowych pokrywa się z ich liczbą, dla innych jest równa iloczynowi liczby więzadeł i ich uzębienia. Charakter L. determinuje rodzaje koordynacji. połączenie (kompleksy aminowe, addukty molowe, chelaty, kompleksy p itp.); od tego zależą właściwości, struktura i reakcja. zdolność do skomplikowanych połączeń i możliwość ich praktycznego zastosowania Aplikacje. Oświetlony.: Garnevsky A. D., „Izwiestia. Wyższe instytucje edukacyjne, seria. Chemia i technologia chemiczna”. 1987, t. 30, t. 10, s. 10 3-16; „Coord.”, 1988, t. 14, ok. 2000-2000. 5, s. 579-99; Kompleksowa chemia koordynacyjna, v. 2-Ligandy, Oxf., 1987. A. D. Garnowski.
Encyklopedia chemiczna. - M .: Encyklopedia radziecka. wyd. I. L. Knunyants. 1988 .
Zobacz, jakie „LIGANDY” znajdują się w innych słownikach:
- (z łac. ligo wiążę) na przykład w złożonych związkach, cząsteczkach lub jonach związanych z atomem centralnym (czynnikiem kompleksującym). w związku LIGATURA (późne łac. połączenie ligatura) 1) litera lub znak transkrypcji fonetycznej, utworzona z ... ... Wielki słownik encyklopedyczny
ligandy- (dodaje) – jony, rodniki lub cząsteczki obojętne, które lokalizują się wokół jonu centralnego (atomu) w wyniku utworzenia wiązania koordynacyjnego. Chemia ogólna: podręcznik / A. V. Zholnin ... Terminy chemiczne
- (od łacińskiego ligo I bind), w związkach złożonych występują cząsteczki lub jony związane z centralnym atomem (czynnikiem kompleksującym), np. w związku Cl3 występuje centralny atom Co, a ligandami są cząsteczki NH3. * * * LIGANDY LIGANDY (od łac. Ligo ... słownik encyklopedyczny
- (z łac. ligo wiążę) w związkach złożonych (patrz Związki złożone) cząsteczki lub jony bezpośrednio związane z atomem centralnym; tak samo jak dodatki... Duży Encyklopedia radziecka
- (od łac. ligo łączę), w skomplikowanych pojemnikach. cząsteczki lub jony związane z centrum, atomem (czynnikiem kompleksującym), np. w połączeniu Centrum [Co(NH3)6]C13. Atom Co i cząsteczka L. NH3... Naturalna nauka. słownik encyklopedyczny
- ... Wikipedii
Ligandy- (łac. ligo – wiążę). Specyficzne substancje endogenne, które pobudzają określone typy komórkowych receptorów mózgowych - adrenergiczne, dopaminergiczne, cholinergiczne, serotoninergiczne, benzodiazepinowe, peptydergiczne itp. Słownik wyjaśniający terminów psychiatrycznych
ligandy- lig andy, ow, jednostki. ręka i... Słownik ortografii rosyjskiej
Ligandy- (łac. ligo bind) substancje endogenne, które są neuroprzekaźnikami w synapsach mózgu (dopamina, acetylocholina, serotonina itp.). Leki psychotropowe i halucynogeny również mają pewne właściwości ligandów. * * * Biologicznie… … Encyklopedyczny słownik psychologii i pedagogiki
ligandy osiowe- ašiniai ligandai statusas T sritis chemija apibrėžtis Vienoje ašyje esantys ligandai. atitikmenys: pol. ligandy osiowe rus. ligandy osiowe ryšiai: sinonimas – aksialiniai ligandai… Chemijos terminų aiškinamasis žodynas
Książki
- Immunologiczne problemy apoptozy, A. Yu. Barysznikow, Yu. V. Shishkin. Ostatnia dekada charakteryzowało się szybkim badaniem procesu programowanej śmierci komórki (apoptozy). Odkryto receptory na powierzchni komórki i ich ligandy, które pośredniczą w...
Struktura modułu | Motywy |
Jednostka modułowa 1 | 1.1. Strukturalna organizacja białek. Etapy tworzenia konformacji białka natywnego 1.2. Podstawy funkcjonowania białek. Leki jako ligandy wpływające na funkcję białek 1.3. Denaturacja białek i możliwość ich samoistnej renatywacji |
Jednostka modułowa 2 | 1.4. Cechy budowy i funkcjonowania białek oligomerycznych na przykładzie hemoglobiny 1,5. Utrzymanie konformacji białka natywnego w warunkach komórkowych 1.6. Różnorodność białek. Rodziny białek na przykładzie immunoglobulin 1.7. Właściwości fizykochemiczne białek i metody ich rozdzielania |
Jednostka modułowa 1 ORGANIZACJA STRUKTURALNA BIAŁEK MONOMERYCZNYCH I PODSTAWY ICH DZIAŁANIA
Cele nauczania Być w stanie:
1. Wykorzystywać wiedzę o cechach strukturalnych białek i zależności funkcji białek od ich budowy do zrozumienia mechanizmów rozwoju proteinopatii dziedzicznych i nabytych.
2. Wyjaśniać mechanizmy działania terapeutycznego niektórych leków jako ligandów oddziałujących z białkami i zmieniających ich działanie.
3. Wykorzystywać wiedzę o strukturze i labilności konformacyjnej białek do zrozumienia ich niestabilności strukturalnej i funkcjonalnej oraz tendencji do denaturacji w zmieniających się warunkach.
4. Wyjaśniać zastosowanie środków denaturujących do sterylizacji materiałów i narzędzi medycznych oraz środków antyseptycznych.
Wiedzieć:
1. Poziomy organizacji strukturalnej białek.
2. Znaczenie struktury pierwszorzędowej białek, która decyduje o ich różnorodności strukturalnej i funkcjonalnej.
3. Mechanizm powstawania centrum aktywnego w białkach i jego specyficzne oddziaływanie z ligandem, które leży u podstaw funkcjonowania białek.
4. Przykłady wpływu egzogennych ligandów (leków, toksyn, trucizn) na konformację i aktywność funkcjonalną białek.
5. Przyczyny i skutki denaturacji białek, czynniki powodujące denaturację.
6. Przykłady zastosowania czynników denaturujących w medycynie jako środków antyseptycznych i środków do sterylizacji narzędzi medycznych.
TEMAT 1.1. ORGANIZACJA STRUKTURALNA BIAŁEK. ETAPY FORMOWANIA RODZIMYCH
KONFORMACJE BIAŁKOWE
Białka to cząsteczki polimerów, których monomerami jest tylko 20 α-aminokwasów. Zestaw i kolejność kombinacji aminokwasów w białku jest zdeterminowana strukturą genów w DNA poszczególnych osobników. Każde białko, zgodnie ze swoją specyficzną budową, pełni swoją funkcję. Zestaw białek danego organizmu decyduje o jego cechach fenotypowych, a także o występowaniu chorób dziedzicznych lub predyspozycji do ich rozwoju.
1. Aminokwasy tworzące białka. Wiązanie peptydowe. Białka to polimery zbudowane z monomerów – 20 α-aminokwasów, których ogólny wzór to
Aminokwasy różnią się budową, wielkością i właściwościami fizykochemicznymi rodników przyłączonych do atomu węgla α. Grupy funkcyjne aminokwasów określają charakterystykę właściwości różnych α-aminokwasów. Rodniki występujące w α-aminokwasach można podzielić na kilka grup:
prolina, W przeciwieństwie do pozostałych 19 monomerów białkowych, nie jest to aminokwas, ale iminokwas; rodnik w prolinie jest związany zarówno z atomem węgla α, jak i grupą iminową
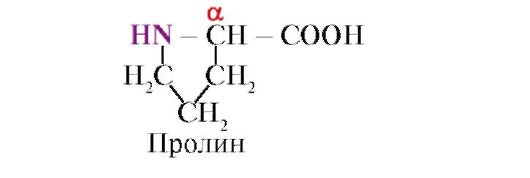 Aminokwasy różnią się rozpuszczalnością w wodzie. Wynika to ze zdolności rodników do interakcji z wodą (hydratem).
Aminokwasy różnią się rozpuszczalnością w wodzie. Wynika to ze zdolności rodników do interakcji z wodą (hydratem).
DO hydrofilowy obejmują rodniki zawierające anionowe, kationowe i polarne nienaładowane grupy funkcyjne.
DO hydrofobowy obejmują rodniki zawierające grupy metylowe, łańcuchy lub pierścienie alifatyczne.
2. Wiązania peptydowe łączą aminokwasy, tworząc peptydy. Podczas syntezy peptydów grupa α-karboksylowa jednego aminokwasu oddziałuje z grupą α-aminową innego aminokwasu, tworząc wiązanie peptydowe:
Białka są polipeptydami, tj. liniowe polimery α-aminokwasów połączone wiązaniem peptydowym (ryc. 1.1.)
 Ryż. 1.1. Terminy używane do opisu struktury peptydów
Ryż. 1.1. Terminy używane do opisu struktury peptydów
Nazywa się monomery aminokwasów tworzące polipeptydy reszty aminokwasowe.Łańcuch powtarzających się grup - NH-CH-CO- formy szkielet peptydowy. Resztę aminokwasu zawierającą wolną grupę α-aminową nazywa się N-końcową, a resztę zawierającą wolną grupę α-karboksylową nazywa się C-końcową. Peptydy są zapisywane i odczytywane od N-końca do C-końca.
Wiązanie peptydowe utworzone przez grupę iminową proliny różni się od innych wiązań peptydowych: atom azotu grupy peptydowej nie zawiera wodoru,
zamiast tego dochodzi do wiązania z rodnikiem, w wyniku czego jedna strona pierścienia zostaje włączona do szkieletu peptydowego:
 Peptydy różnią się składem aminokwasów, liczbą aminokwasów i kolejnością połączeń aminokwasów, na przykład Ser-Ala-Glu-Gis i His-Glu-Ala-Ser to dwa różne peptydy.
Peptydy różnią się składem aminokwasów, liczbą aminokwasów i kolejnością połączeń aminokwasów, na przykład Ser-Ala-Glu-Gis i His-Glu-Ala-Ser to dwa różne peptydy.
Wiązania peptydowe są bardzo mocne, a ich chemiczna, nieenzymatyczna hydroliza wymaga trudnych warunków: analizowane białko poddaje się hydrolizie w stężonym kwasie solnym w temperaturze około 110° przez 24 godziny. W żywej komórce wiązania peptydowe mogą zostać rozerwane Enzymy proteolityczne, zwany proteazy Lub hydrolazy peptydowe.
3. Podstawowa struktura białek. Reszty aminokwasowe w łańcuchach peptydowych różnych białek nie zmieniają się losowo, ale są ułożone w określonej kolejności. Nazywa się sekwencją liniową lub kolejnością naprzemienną reszt aminokwasowych w łańcuchu polipeptydowym pierwotna struktura białka.
Podstawowa struktura każdego pojedynczego białka jest kodowana w cząsteczce DNA (w regionie zwanym genem) i jest realizowana podczas transkrypcji (kopiowanie informacji na mRNA) i translacji (synteza pierwszorzędowej struktury białka). Zatem pierwotną strukturą białek pojedynczego człowieka jest informacja przekazywana dziedzicznie z rodziców na dzieci, która określa cechy strukturalne białek danego organizmu, od których zależy funkcja istniejących białek (ryc. 1.2.).
 Ryż. 1.2. Związek pomiędzy genotypem a konformacją białek syntetyzowanych w organizmie człowieka
Ryż. 1.2. Związek pomiędzy genotypem a konformacją białek syntetyzowanych w organizmie człowieka
Każde z około 100 000 pojedynczych białek w organizmie człowieka ma unikalny struktura pierwotna. Cząsteczki tego samego rodzaju białka (na przykład albuminy) mają tę samą przemianę reszt aminokwasowych, co odróżnia albuminę od innych pojedynczych białek.
Sekwencję reszt aminokwasowych w łańcuchu peptydowym można uznać za formę zapisu informacji. Informacje te określają przestrzenne rozmieszczenie liniowego łańcucha peptydowego w bardziej zwartą trójwymiarową strukturę zwaną struktura wiewiórka. Nazywa się proces tworzenia funkcjonalnie aktywnej konformacji białka składanie
4. Konformacja białka. Swobodna rotacja w szkielecie peptydowym jest możliwa pomiędzy atomem azotu grupy peptydowej a sąsiadującym atomem węgla α, a także pomiędzy atomem węgla α a węglem grupy karbonylowej. Dzięki oddziaływaniu grup funkcyjnych reszt aminokwasowych pierwotna struktura białek może nabrać bardziej złożonych struktur przestrzennych. W białkach globularnych istnieją dwa główne poziomy fałdowania konformacji łańcuchów peptydowych: wtórny I struktura trzeciorzędowa.
Struktura wtórna białek to struktura przestrzenna powstająca w wyniku utworzenia wiązań wodorowych pomiędzy grupami funkcyjnymi -C=O i -NH- szkieletu peptydowego. W tym przypadku łańcuch peptydowy może uzyskać regularne struktury dwóch typów: α-helisy I Struktury β.
W α-helisy wiązania wodorowe powstają między atomem tlenu grupy karbonylowej a wodorem azotu amidowego czwartego aminokwasu; łańcuchy boczne reszt aminokwasowych
znajdują się wzdłuż obwodu spirali, nie uczestnicząc w tworzeniu struktury wtórnej (ryc. 1.3.).
Rodniki masowe lub rodniki niosące równe ładunki zapobiegają tworzeniu się α-helisy. Reszta proliny, która ma strukturę pierścieniową, przerywa α-helisę, ponieważ z powodu braku wodoru przy atomie azotu w łańcuchu peptydowym niemożliwe jest utworzenie wiązania wodorowego. Wiązanie między azotem i atomem węgla α jest częścią pierścienia proliny, więc szkielet peptydowy ulega w tym miejscu wygięciu.
Struktura β powstaje pomiędzy liniowymi regionami szkieletu peptydowego jednego łańcucha polipeptydowego, tworząc w ten sposób złożone struktury. Mogą tworzyć się łańcuchy polipeptydowe lub ich części równoległy Lub antyrównoległe struktury β. W pierwszym przypadku N- i C-końce oddziałujących łańcuchów peptydowych pokrywają się, w drugim mają przeciwny kierunek (ryc. 1.4).
Ryż. 1.3. Struktura drugorzędowa białka - α-helisa
 Ryż. 1.4. Struktury β-arkuszowe równoległe i antyrównoległe
Ryż. 1.4. Struktury β-arkuszowe równoległe i antyrównoległe
Struktury β zaznaczono szerokimi strzałkami: A – Struktura β antyrównoległa. B - Równoległe struktury β-arkuszowe
W niektórych białkach struktury β mogą powstawać w wyniku tworzenia się wiązań wodorowych pomiędzy atomami szkieletu peptydowego różnych łańcuchów polipeptydowych.
Występuje także w białkach obszary o nieregularnym wtórnym struktura, która obejmuje zagięcia, pętle i zwoje szkieletu polipeptydowego. Często lokalizują się w miejscach, w których zmienia się kierunek łańcucha peptydowego, np. gdy tworzy się równoległa struktura β-kartki.
Na podstawie obecności α-helis i β-struktur białka globularne można podzielić na cztery kategorie.
Ryż. 1,5. Wtórna struktura mioglobiny (A) i łańcucha β hemoglobiny (B), zawierająca osiem α-helis

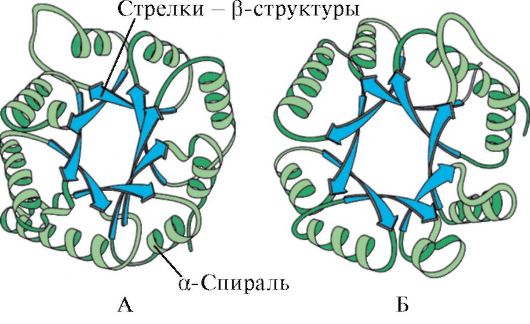 Ryż. 1.6. Struktura drugorzędowa domeny izomerazy triozofosforanowej i kinazy pirogronianowej
Ryż. 1.6. Struktura drugorzędowa domeny izomerazy triozofosforanowej i kinazy pirogronianowej
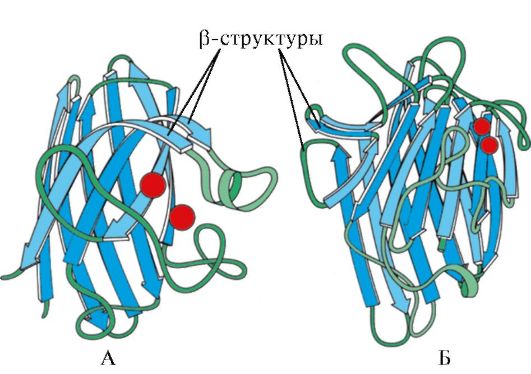 Ryż. 1.7. Struktura drugorzędowa domeny stałej immunoglobuliny (A) i enzymu dysmutazy ponadtlenkowej (B)
Ryż. 1.7. Struktura drugorzędowa domeny stałej immunoglobuliny (A) i enzymu dysmutazy ponadtlenkowej (B)
W czwarta kategoria obejmowały białka zawierające niewielką ilość regularnych struktur drugorzędowych. Białka te obejmują małe białka bogate w cysteinę lub metaloproteiny.
Trzeciorzędowa struktura białka- rodzaj konformacji powstający w wyniku oddziaływań pomiędzy rodnikami aminokwasów, które w łańcuchu peptydowym mogą znajdować się w znacznej odległości od siebie. Większość białek tworzy przestrzenną strukturę przypominającą globulę (białka globularne).
Ponieważ hydrofobowe rodniki aminokwasowe mają tendencję do łączenia się poprzez tzw oddziaływania hydrofobowe i międzycząsteczkowe siły van der Waalsa wewnątrz globulki białkowej tworzy się gęsty hydrofobowy rdzeń. Hydrofilowe rodniki zjonizowane i niezjonizowane zlokalizowane są głównie na powierzchni białka i decydują o jego rozpuszczalności w wodzie.
 Ryż. 1.8. Rodzaje wiązań powstających pomiędzy rodnikami aminokwasów podczas tworzenia trzeciorzędowej struktury białka
Ryż. 1.8. Rodzaje wiązań powstających pomiędzy rodnikami aminokwasów podczas tworzenia trzeciorzędowej struktury białka
1 - wiązanie jonowe- występuje pomiędzy dodatnio i ujemnie naładowanymi grupami funkcyjnymi;
2 - wiązanie wodorowe- występuje pomiędzy hydrofilową grupą nienaładowaną i dowolną inną grupą hydrofilową;
3 - oddziaływania hydrofobowe- powstają pomiędzy rodnikami hydrofobowymi;
4 - wiązanie disiarczkowe- powstają w wyniku utleniania grup SH reszt cysteiny i ich wzajemnego oddziaływania
Hydrofilowe reszty aminokwasowe znajdujące się wewnątrz hydrofobowego rdzenia mogą oddziaływać ze sobą za pomocą joński I wiązania wodorowe(ryc. 1.8).
Wiązania jonowe, wodorowe i oddziaływania hydrofobowe są słabe: ich energia jest niewiele wyższa od energii ruchu termicznego cząsteczek w temperaturze pokojowej. Konformacja białka jest utrzymywana poprzez tworzenie wielu takich słabych wiązań. Ponieważ atomy tworzące białko są w ciągłym ruchu, możliwe jest rozerwanie niektórych słabych wiązań i utworzenie innych, co prowadzi do nieznacznych ruchów poszczególnych odcinków łańcucha polipeptydowego. Nazywa się tę właściwość białek zmiany konformacji w wyniku rozrywania niektórych i tworzenia innych słabych wiązań labilność konformacyjna.
Ciało ludzkie ma systemy, które wspierają homeostaza- stałość środowiska wewnętrznego w pewnych dopuszczalnych granicach dla zdrowego organizmu. W warunkach homeostazy niewielkie zmiany w konformacji nie zakłócają ogólnej struktury i funkcji białek. Nazywa się funkcjonalnie aktywną konformację białka konformacja natywna. Zmiany w środowisku wewnętrznym (na przykład stężenie glukozy, jonów Ca, protonów itp.) prowadzą do zmian w konformacji i zakłócenia funkcji białek.
Trzeciorzędowa struktura niektórych białek jest ustabilizowana wiązania disiarczkowe, powstaje w wyniku oddziaływania grup -SH dwóch reszt
 Ryż. 1.9. Tworzenie wiązania dwusiarczkowego w cząsteczce białka
Ryż. 1.9. Tworzenie wiązania dwusiarczkowego w cząsteczce białka
cysteina (ryc. 1.9). Większość białek wewnątrzkomórkowych nie ma kowalencyjnych wiązań dwusiarczkowych w swojej trzeciorzędowej strukturze. Ich obecność jest charakterystyczna dla białek wydzielanych przez komórkę, co zapewnia ich większą stabilność w warunkach zewnątrzkomórkowych. Zatem wiązania dwusiarczkowe są obecne w cząsteczkach insuliny i immunoglobulin.
Insulina- hormon białkowy syntetyzowany w komórkach β trzustki i wydzielany do krwi w odpowiedzi na wzrost stężenia glukozy we krwi. W strukturze insuliny występują dwa wiązania dwusiarczkowe łączące łańcuchy polipeptydowe A i B oraz jedno wiązanie dwusiarczkowe w obrębie łańcucha A (ryc. 1.10).
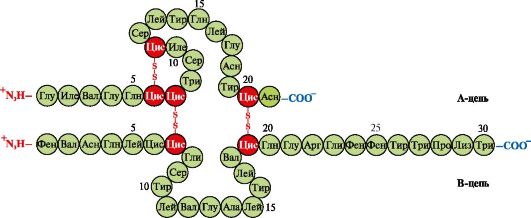 Ryż. 1.10. Wiązania dwusiarczkowe w strukturze insuliny
Ryż. 1.10. Wiązania dwusiarczkowe w strukturze insuliny
5. Nadwtórna struktura białek. Czasami wykrywa się je w białkach o różnej strukturze pierwszorzędowej i funkcjach podobne kombinacje i względne położenia struktur drugorzędnych, które nazywane są strukturą nadwtórną. Zajmuje pozycję pośrednią między strukturami drugorzędowymi i trzeciorzędowymi, ponieważ jest specyficzną kombinacją elementów struktury drugorzędowej w tworzeniu trzeciorzędowej struktury białka. Struktury superwtórne mają specyficzne nazwy, takie jak „α-helisa-zmieniająca helisę”, „zamek leucynowy”, „palce cynkowe” itp. Takie struktury superwtórne są charakterystyczne dla białek wiążących DNA.
„Zamek leucynowy”. Ten typ struktury superwtórnej służy do łączenia ze sobą dwóch białek. Na powierzchni oddziałujących białek znajdują się regiony α-helikalne zawierające co najmniej cztery reszty leucyny. Reszty leucyny w α-helisie są oddalone od siebie o sześć aminokwasów. Ponieważ każdy zwój α-helisy zawiera 3,6 reszt aminokwasowych, rodniki leucyny znajdują się na powierzchni co drugiego zwoju. Reszty leucyny α-helisy jednego białka mogą oddziaływać z resztami leucyny innego białka (oddziaływania hydrofobowe), łącząc je ze sobą (ryc. 1.11.). Wiele białek wiążących DNA funkcjonuje w kompleksach oligomerycznych, w których poszczególne podjednostki są połączone ze sobą „zamkami leucynowymi”.
 Ryż. 1.11. „Zamek leucynowy” pomiędzy regionami α-helikalnymi dwóch białek
Ryż. 1.11. „Zamek leucynowy” pomiędzy regionami α-helikalnymi dwóch białek
Przykładem takich białek są histony. Histony- białka jądrowe, które zawierają dużą liczbę dodatnio naładowanych aminokwasów - argininę i lizynę (do 80%). Cząsteczki histonów łączą się w oligomeryczne kompleksy zawierające osiem monomerów za pomocą „zamków leucynowych”, pomimo znacznego homonimicznego ładunku tych cząsteczek.
„Palec cynkowy”- wariant struktury nadwtórnej, charakterystyczny dla białek wiążących DNA, ma postać wydłużonego fragmentu na powierzchni białka i zawiera około 20 reszt aminokwasowych (ryc. 1.12). Kształt „wydłużonego palca” wspiera atom cynku związany z czterema rodnikami aminokwasowymi – dwiema resztami cysteinowymi i dwiema resztami histydyny. W niektórych przypadkach zamiast reszt histydyny występują reszty cysteiny. Dwie blisko leżące reszty cysteiny są oddzielone od pozostałych dwóch reszt Gisili sekwencją Cys składającą się z około 12 reszt aminokwasowych. Ten region białka tworzy α-helisę, której rodniki mogą specyficznie wiązać się z regionami regulatorowymi głównego rowka DNA. Indywidualna specyficzność wiązania
 Ryż. 1.12. Podstawowa struktura regionu białek wiążących DNA tworzących strukturę „palca cynkowego” (litery wskazują aminokwasy tworzące tę strukturę)
Ryż. 1.12. Podstawowa struktura regionu białek wiążących DNA tworzących strukturę „palca cynkowego” (litery wskazują aminokwasy tworzące tę strukturę)
Białko regulatorowe wiążące DNA zależy od sekwencji reszt aminokwasowych zlokalizowanych w regionie palca cynkowego. Struktury takie zawierają w szczególności receptory dla hormonów steroidowych biorące udział w regulacji transkrypcji (odczycie informacji z DNA na RNA).
TEMAT 1.2. PODSTAWY DZIAŁANIA BIAŁEK. LEKI JAKO LIGANDY WPŁYWAJĄCE NA FUNKCJĘ BIAŁKA
1. Centrum aktywne białka i jego oddziaływanie z ligandem. Podczas tworzenia struktury trzeciorzędowej na powierzchni funkcjonalnie aktywnego białka tworzy się region, zwykle we wnęce utworzonej przez rodniki aminokwasów, które są daleko od siebie oddalone w strukturze pierwszorzędowej. Region ten, który ma unikalną dla danego białka strukturę i jest zdolny do specyficznego oddziaływania z konkretną cząsteczką lub grupą podobnych cząsteczek, nazywany jest miejscem wiązania białko-ligand lub miejscem aktywnym. Ligandy to cząsteczki oddziałujące z białkami.
Wysoka specyficzność Oddziaływanie białka z ligandem zapewnia komplementarność struktury centrum aktywnego do struktury liganda.
Komplementarność- jest to zgodność przestrzenna i chemiczna oddziałujących powierzchni. Centrum aktywne musi nie tylko odpowiadać przestrzennie zawartemu w nim ligandowi, ale także muszą powstać wiązania (oddziaływania jonowe, wodorowe i hydrofobowe) pomiędzy grupami funkcyjnymi rodników wchodzących w skład centrum aktywnego a ligandem, które utrzymują ligand w centrum aktywnym (ryc. 1.13 ).
 Ryż. 1.13. Uzupełniające oddziaływanie białka z ligandem
Ryż. 1.13. Uzupełniające oddziaływanie białka z ligandem
Niektóre ligandy po przyłączeniu do centrum aktywnego białka pełnią pomocniczą rolę w funkcjonowaniu białek. Takie ligandy nazywane są kofaktorami, a białka zawierające część niebiałkową złożone białka(w przeciwieństwie do białek prostych, składających się wyłącznie z części białkowej). Część niebiałkowa, trwale połączona z białkiem, nazywa się grupa protetyczna. Na przykład mioglobina, hemoglobina i cytochromy zawierają grupę prostetyczną, hem, zawierającą jon żelaza, mocno związaną z centrum aktywnym. Złożone białka zawierające hem nazywane są hemoproteinami.
Kiedy do białek przyłączają się specyficzne ligandy, manifestuje się funkcja tych białek. Zatem albumina, najważniejsze białko osocza krwi, spełnia swoją funkcję transportową poprzez przyłączenie do centrum aktywnego ligandów hydrofobowych, takich jak kwasy tłuszczowe, bilirubina, niektóre leki itp. (ryc. 1.14)
Ligandami oddziałującymi z trójwymiarową strukturą łańcucha peptydowego mogą być nie tylko drobnocząsteczkowe cząsteczki organiczne i nieorganiczne, ale także makrocząsteczki:
DNA (przykłady z białkami wiążącymi DNA omówione powyżej);
Polisacharydy;
 Ryż. 1.14. Związek między genotypem a fenotypem
Ryż. 1.14. Związek między genotypem a fenotypem
Unikalna pierwotna struktura białek ludzkich, zakodowana w cząsteczce DNA, realizowana jest w komórkach w postaci unikalnej konformacji, struktury centrum aktywnego i funkcji białek
W takich przypadkach białko rozpoznaje specyficzny region ligandu, który jest proporcjonalny i komplementarny do miejsca wiązania. Zatem na powierzchni hepatocytów znajdują się białka receptorowe dla hormonu insuliny, który również ma strukturę białkową. Oddziaływanie insuliny z receptorem powoduje zmianę jej konformacji i aktywację układów sygnalizacyjnych, co prowadzi do magazynowania składników odżywczych w hepatocytach po posiłkach.
Zatem, Funkcjonowanie białek opiera się na specyficznym oddziaływaniu centrum aktywnego białka z ligandem.
2. Struktura domeny i jej rola w funkcjonowaniu białek. Długie łańcuchy polipeptydowe białek globularnych często składają się na kilka zwartych, stosunkowo niezależnych regionów. Mają niezależną strukturę trzeciorzędową, przypominającą białka globularne i nazywane są domeny. Dzięki strukturze domenowej białek łatwiej jest uformować ich strukturę trzeciorzędową.
W białkach domenowych miejsca wiązania ligandów często znajdują się pomiędzy domenami. Zatem trypsyna jest enzymem proteolitycznym wytwarzanym przez zewnątrzwydzielniczą część trzustki i jest niezbędna do trawienia białek pokarmowych. Ma budowę dwudomenową, a centrum wiązania trypsyny z jej ligandem – białkiem pokarmowym – znajduje się w rowku pomiędzy obiema domenami. W centrum aktywnym powstają warunki niezbędne do skutecznego wiązania określonego miejsca białka spożywczego i hydrolizy jego wiązań peptydowych.
Różne domeny w białku mogą przemieszczać się względem siebie, gdy centrum aktywne oddziałuje z ligandem (ryc. 1.15).
Heksokinaza- enzym katalizujący fosforylację glukozy przy użyciu ATP. Miejsce aktywne enzymu znajduje się w szczelinie pomiędzy dwiema domenami. Kiedy heksokinaza wiąże się z glukozą, otaczające ją domeny zamykają się, a substrat zostaje uwięziony, gdzie następuje fosforylacja (patrz ryc. 1.15).
 Ryż. 1,15. Wiązanie domen heksokinazy z glukozą
Ryż. 1,15. Wiązanie domen heksokinazy z glukozą
W niektórych białkach domeny pełnią niezależne funkcje, wiążąc się z różnymi ligandami. Takie białka nazywane są wielofunkcyjnymi.
3. Leki to ligandy wpływające na funkcję białek. Oddziaływanie białek z ligandami jest specyficzne. Jednakże, ze względu na labilność konformacyjną białka i jego centrum aktywnego, możliwe jest wybranie innej substancji, która również mogłaby oddziaływać z białkiem w centrum aktywnym lub innej części cząsteczki.
Nazywa się substancję o strukturze podobnej do naturalnego ligandu Strukturalny analog ligandu lub nienaturalny ligand. Oddziałuje również z białkiem w miejscu aktywnym. Strukturalny analog liganda może zarówno wzmacniać funkcję białka (agonista), i zmniejsz go (antagonista). Ligand i jego analogi strukturalne konkurują ze sobą o wiązanie z białkiem w tym samym miejscu. Takie substancje nazywane są konkurencyjne modulatory(regulatory) funkcji białek. Wiele leków działa jako inhibitory białek. Część z nich otrzymywana jest poprzez chemiczną modyfikację naturalnych ligandów. Inhibitorami funkcji białek mogą być leki i trucizny.
Atropina jest konkurencyjnym inhibitorem receptorów M-cholinergicznych. Acetylocholina jest neuroprzekaźnikiem odpowiedzialnym za przekazywanie impulsów nerwowych przez synapsy cholinergiczne. Aby przeprowadzić wzbudzenie, acetylocholina uwolniona do szczeliny synaptycznej musi oddziaływać z białkiem receptorowym błony postsynaptycznej. Znaleziono dwa typy receptory cholinergiczne:
Receptor M oprócz acetylocholiny oddziałuje selektywnie z muskaryną (toksyną muchomora). M - receptory cholinergiczne występują na mięśniach gładkich i podczas interakcji z acetylocholiną powodują ich skurcz;
Receptor H specyficznie wiążący się z nikotyną. Receptory N-cholinergiczne znajdują się w synapsach prążkowanych mięśni szkieletowych.
Specyficzny inhibitor Receptory M-cholinergiczne jest atropina. Występuje w roślinach belladonna i lulek.
 Atropina posiada grupy funkcyjne podobne budową do acetylocholiny i ich rozmieszczeniem przestrzennym, dlatego jest konkurencyjnym inhibitorem receptorów M-cholinergicznych. Biorąc pod uwagę, że wiązanie acetylocholiny z receptorami M-cholinergicznymi powoduje skurcz mięśni gładkich, atropina jest stosowana jako lek łagodzący ich skurcze (przeciwskurczowy). Dlatego wiadomo, że atropina służy do rozluźniania mięśni oka podczas oglądania dna oka, a także do łagodzenia skurczów podczas kolki żołądkowo-jelitowej. Receptory M-cholinergiczne są również obecne w ośrodku system nerwowy(OUN), dlatego duże dawki atropiny mogą powodować niepożądane reakcje ze strony ośrodkowego układu nerwowego: pobudzenie ruchowe i psychiczne, omamy, drgawki.
Atropina posiada grupy funkcyjne podobne budową do acetylocholiny i ich rozmieszczeniem przestrzennym, dlatego jest konkurencyjnym inhibitorem receptorów M-cholinergicznych. Biorąc pod uwagę, że wiązanie acetylocholiny z receptorami M-cholinergicznymi powoduje skurcz mięśni gładkich, atropina jest stosowana jako lek łagodzący ich skurcze (przeciwskurczowy). Dlatego wiadomo, że atropina służy do rozluźniania mięśni oka podczas oglądania dna oka, a także do łagodzenia skurczów podczas kolki żołądkowo-jelitowej. Receptory M-cholinergiczne są również obecne w ośrodku system nerwowy(OUN), dlatego duże dawki atropiny mogą powodować niepożądane reakcje ze strony ośrodkowego układu nerwowego: pobudzenie ruchowe i psychiczne, omamy, drgawki.
Ditilin jest konkurencyjnym agonistą receptorów H-cholinergicznych, hamującym funkcję synaps nerwowo-mięśniowych.
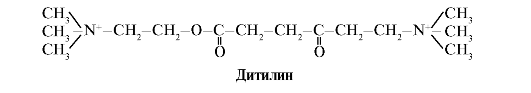 Synapsy nerwowo-mięśniowe mięśni szkieletowych zawierają receptory H-cholinergiczne. Ich interakcja z acetylocholiną prowadzi do skurczów mięśni. Podczas niektórych zabiegów chirurgicznych, a także w badaniach endoskopowych stosuje się leki powodujące rozluźnienie mięśni szkieletowych (leki zwiotczające mięśnie). Należą do nich ditilina, która jest strukturalnym analogiem acetylocholiny. Przyłącza się do receptorów H-cholinergicznych, ale w przeciwieństwie do acetylocholiny jest bardzo powoli niszczona przez enzym acetylocholinoesterazę. W wyniku długotrwałego otwarcia kanałów jonowych i utrzymującej się depolaryzacji błony dochodzi do zaburzenia przewodzenia impulsów nerwowych i rozluźnienia mięśni. Początkowo właściwości te odkryto w truciźnie kurary, dlatego nazywane są takimi lekami podobny do kurary.
Synapsy nerwowo-mięśniowe mięśni szkieletowych zawierają receptory H-cholinergiczne. Ich interakcja z acetylocholiną prowadzi do skurczów mięśni. Podczas niektórych zabiegów chirurgicznych, a także w badaniach endoskopowych stosuje się leki powodujące rozluźnienie mięśni szkieletowych (leki zwiotczające mięśnie). Należą do nich ditilina, która jest strukturalnym analogiem acetylocholiny. Przyłącza się do receptorów H-cholinergicznych, ale w przeciwieństwie do acetylocholiny jest bardzo powoli niszczona przez enzym acetylocholinoesterazę. W wyniku długotrwałego otwarcia kanałów jonowych i utrzymującej się depolaryzacji błony dochodzi do zaburzenia przewodzenia impulsów nerwowych i rozluźnienia mięśni. Początkowo właściwości te odkryto w truciźnie kurary, dlatego nazywane są takimi lekami podobny do kurary.
TEMAT 1.3. DENATURACJA BIAŁEK I MOŻLIWOŚĆ ICH SPONTANICZNEJ RENATURACJI
1. Ponieważ natywna konformacja białek jest utrzymywana na skutek słabych oddziaływań, zmiany składu i właściwości środowiska otaczającego białko, narażenie na odczynniki chemiczne i czynniki fizyczne powodują zmianę ich konformacji (właściwość labilności konformacyjnej). Zerwanie dużej liczby wiązań prowadzi do zniszczenia natywnej konformacji i denaturacji białek.
Denaturacja białek- jest to zniszczenie ich natywnej konformacji pod wpływem czynników denaturujących, spowodowane zerwaniem słabych wiązań stabilizujących strukturę przestrzenną białka. Denaturacji towarzyszy zniszczenie unikalnej trójwymiarowej struktury i centrum aktywnego białka oraz utrata jego aktywności biologicznej (ryc. 1.16).
Wszystkie zdenaturowane cząsteczki jednego białka uzyskują losową konformację, która różni się od innych cząsteczek tego samego białka. Rodniki aminokwasowe tworzące centrum aktywne okazują się być od siebie przestrzennie odległe, tj. specyficzne miejsce wiązania białka z ligandem ulega zniszczeniu. Podczas denaturacji pierwotna struktura białek pozostaje niezmieniona.
Zastosowanie środków denaturujących w badaniach biologicznych i medycynie. W badaniach biochemicznych przed oznaczeniem w materiale biologicznym związków o niskiej masie cząsteczkowej zazwyczaj najpierw usuwa się z roztworu białka. Najczęściej wykorzystuje się w tym celu kwas trichlorooctowy (TCA). Po dodaniu TCA do roztworu zdenaturowane białka wytrącają się i można je łatwo usunąć przez filtrację (tab. 1.1.).
W medycynie środki denaturujące są często stosowane do sterylizacji narzędzi i materiałów medycznych w autoklawach (środkiem denaturującym jest wysoka temperatura) oraz jako środki antyseptyczne (alkohol, fenol, chloramina) do leczenia zanieczyszczonych powierzchni zawierających patogenną mikroflorę.
2. Spontaniczna reaktywacja białek- dowód determinizmu pierwotnej struktury, konformacji i funkcji białek. Poszczególne białka są produktami jednego genu, które mają identyczną sekwencję aminokwasów i uzyskują w komórce tę samą konformację. Zasadniczy wniosek, że pierwotna struktura białka zawiera już informację o jego konformacji i funkcji, został wysunięty na podstawie zdolności niektórych białek (w szczególności rybonukleazy i mioglobiny) do samoistnej renatywacji – przywracania ich natywnej konformacji po denaturacji.
Tworzenie przestrzennych struktur białkowych odbywa się metodą samoorganizacji – spontanicznego procesu, w którym łańcuch polipeptydowy, posiadający unikalną strukturę pierwotną, ma tendencję do przyjmowania konformacji o najniższej energii swobodnej w roztworze. Zdolność do renatywacji białek, które po denaturacji zachowują swoją pierwotną strukturę, opisano w eksperymencie z enzymem rybonukleazą.
Rybonukleaza jest enzymem rozkładającym wiązania pomiędzy poszczególnymi nukleotydami w cząsteczce RNA. To globularne białko ma jeden łańcuch polipeptydowy, którego trzeciorzędowa struktura jest stabilizowana przez wiele słabych i cztery wiązania dwusiarczkowe.
Traktowanie rybonukleazy mocznikiem, który rozrywa wiązania wodorowe w cząsteczce, oraz czynnikiem redukującym, który rozrywa wiązania disiarczkowe, prowadzi do denaturacji enzymu i utraty jego aktywności.
Usunięcie czynników denaturujących poprzez dializę prowadzi do przywrócenia konformacji i funkcji białka, tj. do odrodzenia. (ryc. 1.17).
 Ryż. 1.17. Denaturacja i renatywacja rybonukleazy
Ryż. 1.17. Denaturacja i renatywacja rybonukleazy
A - natywna konformacja rybonukleazy, w której trzeciorzędowej strukturze znajdują się cztery wiązania dwusiarczkowe; B - zdenaturowana cząsteczka rybonukleazy;
B - reaktywowana cząsteczka rybonukleazy z przywróconą strukturą i funkcją
1. Wypełnij tabelę 1.2.
Tabela 1.2. Klasyfikacja aminokwasów ze względu na polarność rodników
2. Zapisz wzór tetrapeptydu:
Asp – Pro – Fen – Liz
a) podkreślić powtarzające się grupy w peptydzie, które tworzą szkielet peptydu i grupy zmienne reprezentowane przez rodniki aminokwasowe;
b) oznaczyć końce N i C;
c) podkreślić wiązania peptydowe;
d) napisz kolejny peptyd składający się z tych samych aminokwasów;
d) policzyć ilość możliwe opcje tetrapeptyd o podobnym składzie aminokwasów.
3. Wyjaśnij rolę pierwszorzędowej struktury białek na przykładzie analizy porównawczej dwóch strukturalnie podobnych i bliskich ewolucyjnie hormonów peptydowych neuroprzysadki ssaków – oksytocyny i wazopresyny (tab. 1.3).
Tabela 1.3. Struktura i funkcje oksytocyny i wazopresyny
 Dla tego:
Dla tego:
a) porównać skład i sekwencję aminokwasów dwóch peptydów;
b) znaleźć podobieństwo struktury pierwszorzędowej obu peptydów i podobieństwo ich działania biologicznego;
c) znaleźć różnice w budowie dwóch peptydów i różnice w ich funkcjach;
d) wyciągnąć wniosek na temat wpływu struktury pierwszorzędowej peptydów na ich funkcje.
4. Opisać główne etapy powstawania konformacji białek globularnych (struktury drugorzędowe, trzeciorzędowe, koncepcja struktury superwtórnej). Wskaż rodzaje wiązań biorących udział w tworzeniu struktur białkowych. Które rodniki aminokwasowe mogą brać udział w tworzeniu oddziaływań hydrofobowych, jonowych, wiązań wodorowych.
Daj przykłady.
5. Zdefiniować pojęcie „labilności konformacyjnej białek”, wskazać przyczyny jego istnienia i znaczenie.
6. Rozwiń znaczenie wyrażenia: „Funkcjonowanie białek opiera się na ich specyficznej interakcji z ligandem”, używając terminów i wyjaśniając ich znaczenie: konformacja białka, centrum aktywne, ligand, komplementarność, funkcja białka.
7. Na jednym przykładzie wyjaśnij, czym są domeny i jaka jest ich rola w funkcjonowaniu białek.
ZADANIA SAMOKONTROLI
1. Mecz.
Grupa funkcyjna rodnika aminokwasowego:
A. Grupa karboksylowa B. Grupa hydroksylowa C Grupa guanidynowa D. Grupa tiolowa E. Grupa aminowa
2. Wybierz prawidłowe odpowiedzi.
Aminokwasy z polarnymi nienaładowanymi rodnikami to:
A. Cis B. Asn
B. Glu G. Trzy
3. Wybierz prawidłowe odpowiedzi.
Rodniki aminokwasowe:
A. Podaj specyfikę struktury pierwotnej. B. Weź udział w tworzeniu struktury trzeciorzędowej
B. Znajdujące się na powierzchni białka wpływają na jego rozpuszczalność. D. Tworzą centrum aktywne
D. Uczestniczyć w tworzeniu wiązań peptydowych
4. Wybierz prawidłowe odpowiedzi.
Oddziaływania hydrofobowe mogą tworzyć się pomiędzy rodnikami aminokwasów:
A. Tre Lay B. Pro Three
B. Met Ile G. Tir Ala D. Val Fen
5. Wybierz prawidłowe odpowiedzi.
Wiązania jonowe mogą tworzyć się pomiędzy rodnikami aminokwasów:
A. Gln Asp B. Apr Liz
B. Liz Glu G. Gis Asp D. Asn Apr
6. Wybierz prawidłowe odpowiedzi.
Wiązania wodorowe mogą tworzyć się pomiędzy rodnikami aminokwasów:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Mecz.
Rodzaj wiązania biorącego udział w tworzeniu struktury białka:
A. Struktura pierwotna B. Struktura wtórna
B. Struktura trzeciorzędowa
D. Struktura nadwtórna. E. Konformacja.
1. Wiązania wodorowe pomiędzy atomami szkieletu peptydowego
2. Słabe wiązania pomiędzy grupami funkcyjnymi rodników aminokwasów
3. Wiązania pomiędzy grupami α-aminowymi i α-karboksylowymi aminokwasów
8. Wybierz prawidłowe odpowiedzi. Trypsyna:
A. Enzym proteolityczny B. Zawiera dwie domeny
B. Hydrolizuje skrobię
D. Aktywna witryna znajduje się pomiędzy domenami. D. Składa się z dwóch łańcuchów polipeptydowych.
9. Wybierz prawidłowe odpowiedzi. Atropina:
A. Neuroprzekaźnik
B. Strukturalny analog acetylocholiny
B. Oddziałuje z receptorami H-cholinergicznymi
D. Wzmacnia przewodzenie impulsów nerwowych przez synapsy cholinergiczne
D. Konkurencyjny inhibitor receptorów M-cholinergicznych
10. Wybierz poprawne stwierdzenia. W białkach:
A. Struktura pierwotna zawiera informacje o strukturze jej miejsca aktywnego
B. Centrum aktywne powstaje na poziomie struktury pierwotnej
B. Konformacja jest sztywno utwierdzona wiązaniami kowalencyjnymi
D. Miejsce aktywne może oddziaływać z grupą podobnych ligandów
ze względu na labilność konformacyjną białek D. Zmiany w środowisku mogą wpływać na powinowactwo substancji czynnej
centrum do liganda
1. 1-B, 2-G, 3-B.
3. A, B, C, D.
7. 1-B, 2-D, 3-A.
8. A, B, C, D.
PODSTAWOWE TERMINY I POJĘCIA
1. Białko, polipeptyd, aminokwasy
2. Pierwotne, drugorzędowe i trzeciorzędowe struktury białkowe
3. Konformacja, konformacja białka natywnego
4. Wiązania kowalencyjne i słabe w białkach
5. Labilność konformacyjna
6. Miejsce aktywne białka
7. Ligandy
8. Zwijanie białek
9. Strukturalne analogi ligandów
10. Białka domenowe
11. Białka proste i złożone
12. Denaturacja białek, środki denaturujące
13. Reaktywacja białek
Rozwiązywać problemy
„Strukturalna organizacja białek i podstawy ich funkcjonowania”
1. Główną funkcją białka – hemoglobiny A (HbA) jest transport tlenu do tkanek. W populacji ludzkiej znanych jest wiele form tego białka o zmienionych właściwościach i funkcjach – tzw. nieprawidłowe hemoglobiny. Na przykład stwierdzono, że hemoglobina S, występująca w czerwonych krwinkach pacjentów z niedokrwistością sierpowatokrwinkową (HbS), ma słabą rozpuszczalność w warunkach niskiego ciśnienia parcjalnego tlenu (jak ma to miejsce w przypadku krwi żylnej). Prowadzi to do powstania agregatów tego białka. Białko traci swoją funkcję, wytrąca się, a czerwone krwinki przyjmują nieregularny kształt (niektóre z nich przyjmują kształt sierpa) i ulegają szybszemu niż zwykle zniszczeniu w śledzionie. W rezultacie rozwija się anemia sierpowatokrwinkowa.
Jedyną różnicę w pierwszorzędowej strukturze HbA stwierdzono w N-końcowym regionie łańcucha β hemoglobiny. Porównaj regiony N-końcowe nici β i pokaż, jak zmiany w strukturze pierwszorzędowej białka wpływają na jego właściwości i funkcje.
 Dla tego:
Dla tego:
a) napisz wzory aminokwasów, którymi różni się HbA i porównaj właściwości tych aminokwasów (polarność, ładunek).
b) wyciągnąć wniosek co do przyczyny zmniejszenia rozpuszczalności i zakłócenia transportu tlenu do tkanek.
2. Rysunek przedstawia schemat budowy białka, które posiada centrum wiążące z ligandem (centrum aktywne). Wyjaśnij, dlaczego białko dokonuje selektywnego wyboru ligandu. Dla tego:
a) przypomnij sobie, czym jest centrum aktywne białka i rozważ budowę centrum aktywnego białka pokazanego na rysunku;
b) napisz wzory rodników aminokwasowych tworzących centrum aktywne;
c) narysuj ligand, który mógłby specyficznie oddziaływać z miejscem aktywnym białka. Wskaż na nim grupy funkcyjne, które mogą tworzyć wiązania z rodnikami aminokwasów tworzącymi centrum aktywne;
d) wskazać rodzaje wiązań, które powstają pomiędzy ligandem i rodnikami aminokwasowymi centrum aktywnego;
e) wyjaśnić, na czym polega specyfika oddziaływania białko-ligand.
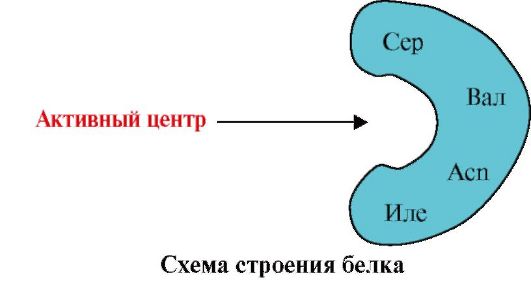 3.
Rysunek przedstawia miejsce aktywne białka i kilka ligandów.
3.
Rysunek przedstawia miejsce aktywne białka i kilka ligandów.
Określ, który ligand najprawdopodobniej oddziałuje z miejscem aktywnym białka i dlaczego.
 Jakie rodzaje wiązań powstają podczas tworzenia kompleksu białko-ligand?
Jakie rodzaje wiązań powstają podczas tworzenia kompleksu białko-ligand?
4. Strukturalne analogi naturalnych ligandów białkowych można stosować jako leki modyfikujące aktywność białek.
Acetylocholina jest mediatorem przekazywania pobudzenia w synapsach nerwowo-mięśniowych. Kiedy acetylocholina wchodzi w interakcję z białkami - receptorami błony postsynaptycznej mięśni szkieletowych, otwierają się kanały jonowe i następuje skurcz mięśni. Ditilin to lek stosowany w niektórych operacjach w celu rozluźnienia mięśni, ponieważ zakłóca przekazywanie impulsów nerwowych przez synapsy nerwowo-mięśniowe. Wyjaśnij mechanizm działania ditiliny jako środka zwiotczającego mięśnie. Dla tego:
a) napisz wzory acetylocholiny i dityliny i porównaj ich budowę;
b) opisać mechanizm relaksującego działania ditiliny.
5. W niektórych chorobach wzrasta temperatura ciała pacjenta, co uważa się za reakcję obronną organizmu. Jednak wysokie temperatury są szkodliwe dla białek organizmu. Wyjaśnij, dlaczego w temperaturach powyżej 40°C funkcja białek zostaje zakłócona i powstaje zagrożenie dla życia człowieka. Aby to zrobić, pamiętaj:
1) Struktura białek i wiązania utrzymujące ich strukturę w natywnej konformacji;
2) Jak zmienia się struktura i funkcja białek wraz ze wzrostem temperatury?;
3) Czym jest homeostaza i dlaczego jest ważna dla utrzymania zdrowia człowieka.
Jednostka modułowa 2 BIAŁKA OLIGOMERYCZNE JAKO CELE DZIAŁAŃ REGULACYJNYCH. STRUKTURALNA I FUNKCJONALNA RÓŻNORODNOŚĆ BIAŁEK. METODY SEPARACJI I OCZYSZCZANIA BIAŁEK
Cele nauczania Być w stanie:
1. Wykorzystywać wiedzę o cechach budowy i funkcjach białek oligomerycznych do zrozumienia adaptacyjnych mechanizmów regulacji ich funkcji.
2. Wyjaśniać rolę białek opiekuńczych w syntezie i utrzymaniu konformacji białek w warunkach komórkowych.
3. Wyjaśniać różnorodność przejawów życia poprzez różnorodność struktur i funkcji białek syntetyzowanych w organizmie.
4. Analizować związek pomiędzy strukturą białek a ich funkcją na przykładach porównania pokrewnych hemoprotein – mioglobiny i hemoglobiny, a także przedstawicieli pięciu klas białek z rodziny immunoglobulin.
5. Wykorzystać wiedzę o osobliwościach właściwości fizycznych i chemicznych białek do doboru metod ich oczyszczania z innych białek i zanieczyszczeń.
6. Interpretować wyniki badań ilościowych i jakość składu białek osocza krwi w celu potwierdzenia lub wyjaśnienia diagnozy klinicznej.
Wiedzieć:
1. Cechy budowy białek oligomerycznych i mechanizmy adaptacyjne regulujące ich funkcje na przykładzie hemoglobiny.
2. Struktura i funkcje białek opiekuńczych oraz ich znaczenie dla utrzymania natywnej konformacji białek w warunkach komórkowych.
3. Zasady łączenia białek w rodziny na podstawie podobieństwa ich budowy i funkcji na przykładzie immunoglobulin.
4. Metody rozdziału białek w oparciu o charakterystykę ich właściwości fizykochemicznych.
5. Elektroforeza osocza krwi jako metoda oceny jakościowego i ilościowego składu białek.
TEMAT 1.4. CECHY STRUKTURY I FUNKCJONOWANIA BIAŁEK OLIGOMERYCZNYCH NA PRZYKŁADZIE HEMOGLOBINY
1. Wiele białek zawiera kilka łańcuchów polipeptydowych. Takie białka nazywane są oligomeryczny, i poszczególne łańcuchy - protomery. Protomery w białkach oligomerycznych są połączone wieloma słabymi wiązaniami niekowalencyjnymi (hydrofobowymi, jonowymi, wodorowymi). Interakcja
protomery realizowane są dzięki komplementarność ich powierzchnie stykowe.
Liczba protomerów w białkach oligomerycznych może się znacznie różnić: hemoglobina zawiera 4 protomery, enzym aminotransferaza asparaginianowa ma 12 protomerów, a białko wirusa mozaiki tytoniu zawiera 2120 protomerów połączonych wiązaniami niekowalencyjnymi. W konsekwencji białka oligomeryczne mogą mieć bardzo wysokie masy cząsteczkowe.
Oddziaływanie jednego protomera z innymi można uznać za szczególny przypadek interakcji białko-ligand, ponieważ każdy protomer służy jako ligand dla innych protomerów. Nazywa się liczbę i sposób łączenia protomerów w białku czwartorzędowa struktura białka.
Białka mogą zawierać protomery o tej samej lub różnej strukturze, na przykład homodimery to białka zawierające dwa identyczne protomery, a heterodimery to białka zawierające dwa różne protomery.
Jeśli białka zawierają różne protomery, wówczas mogą powstać na nich centra wiążące z różnymi ligandami różniącymi się budową. Kiedy ligand wiąże się z miejscem aktywnym, manifestuje się funkcja tego białka. Centrum znajdujące się na innym protomerze nazywa się allosterycznym (innym niż centrum aktywne). Kontaktowanie się allosteryczny ligand lub efektor, pełni funkcję regulacyjną (ryc. 1.18). Oddziaływanie centrum allosterycznego z efektorem powoduje zmiany konformacyjne w strukturze całego białka oligomerycznego ze względu na jego labilność konformacyjną. Wpływa to na powinowactwo miejsca aktywnego do określonego ligandu i reguluje funkcję tego białka. Zmiana konformacji i funkcji wszystkich protomerów podczas interakcji białka oligomerycznego z co najmniej jednym ligandem nazywana jest kooperacyjnymi zmianami konformacyjnymi. Nazywa się efektory wzmacniające funkcję białka aktywatory, i efektory, które hamują jego funkcję - inhibitory.
Zatem białka oligomeryczne, a także białka o strukturze domenowej, mają nową właściwość w porównaniu do białek monomerycznych - zdolność do allosterycznej regulacji funkcji (regulacja poprzez przyłączenie różnych ligandów do białka). Można to zobaczyć porównując struktury i funkcje dwóch blisko spokrewnionych białek złożonych, mioglobiny i hemoglobiny.
 Ryż. 1.18. Schemat struktury białka dimerycznego
Ryż. 1.18. Schemat struktury białka dimerycznego
2. Tworzenie struktur przestrzennych i funkcjonowanie mioglobiny.
Mioglobina (Mb) to białko występujące w mięśniach czerwonych, którego główną funkcją jest tworzenie rezerw O 2 niezbędnych do intensywnej pracy mięśni. Mb jest białkiem złożonym, zawierającym część białkową – apoMb i część niebiałkową – hem. Pierwotna struktura apoMB determinuje jego zwartą konformację kulistą i strukturę centrum aktywnego, do którego przyłączona jest niebiałkowa część mioglobiny – hem. Tlen docierający z krwi do mięśni wiąże się z hemami Fe+ 2 w mioglobinie. Mb jest białkiem monomerycznym, które ma bardzo duże powinowactwo do O 2, dlatego uwalnianie tlenu przez mioglobinę następuje tylko podczas intensywnej pracy mięśni, kiedy ciśnienie parcjalne O 2 gwałtownie maleje.
Tworzenie konformacji Mv. W mięśniach czerwonych na rybosomach podczas translacji syntetyzowana jest pierwotna struktura MB, reprezentowana przez specyficzną sekwencję 153 reszt aminokwasowych. Wtórna struktura Mb zawiera osiem α-helis, zwanych łacińskimi literami od A do H, pomiędzy którymi znajdują się obszary niehelikalne. Trzeciorzędowa struktura Mb ma postać zwartej globuli, w której wnęce znajduje się centrum aktywne pomiędzy α-helisami F i E (ryc. 1.19).
 Ryż. 1.19. Struktura mioglobiny
Ryż. 1.19. Struktura mioglobiny
3. Cechy budowy i funkcjonowania ośrodka czynnego SN. Centrum aktywne Mb utworzone jest głównie przez hydrofobowe rodniki aminokwasowe, szeroko od siebie oddalone w strukturze pierwszorzędowej (na przykład Tri 3 9 i Fen 138) Słabo rozpuszczalne w wodzie ligandy – hem i O 2 – przyłączają się do centrum aktywnego. Hem jest specyficznym ligandem apoMB (ryc. 1.20), którego podstawę stanowią cztery pierścienie pirolowe połączone mostkami metenylowymi; w centrum znajduje się atom Fe+ 2 połączony z atomami azotu pierścieni pirolu czterema wiązaniami koordynacyjnymi. W centrum aktywnym Mb oprócz hydrofobowych rodników aminokwasowych znajdują się także reszty dwóch aminokwasów z rodnikami hydrofilowymi - GisE 7(Rdz 64) i GIS F8(Jego 93) (ryc. 1.21).
 Ryż. 1,20. Struktura hemu - niebiałkowej części mioglobiny i hemoglobiny
Ryż. 1,20. Struktura hemu - niebiałkowej części mioglobiny i hemoglobiny
 Ryż. 1.21. Lokalizacja hemu i O2 w miejscu aktywnym protomerów apomioglobiny i hemoglobiny
Ryż. 1.21. Lokalizacja hemu i O2 w miejscu aktywnym protomerów apomioglobiny i hemoglobiny
Hem jest kowalencyjnie związany z His F8 poprzez atom żelaza. O 2 przyłącza się do żelaza po drugiej stronie płaszczyzny hemu. Jego E 7 jest niezbędny do prawidłowej orientacji O 2 i ułatwia dodanie tlenu do hemu Fe + 2
GIS F8 tworzy wiązanie koordynacyjne z Fe+ 2 i mocno wiąże hem w centrum aktywnym. GisE 7 niezbędny do prawidłowej orientacji w centrum aktywnym innego liganda - O 2 podczas jego interakcji z hemem Fe + 2. Mikrośrodowisko hemu stwarza warunki do silnego, ale odwracalnego wiązania O 2 z Fe + 2 i zapobiega przedostawaniu się wody do hydrofobowego miejsca aktywnego, co może prowadzić do jego utlenienia do Fe + 3.
Monomeryczna struktura Mb i jego centrum aktywnego decyduje o wysokim powinowactwie białka do O2.
4. Struktura oligomeryczna Hb i regulacja powinowactwa Hb do ligandów O 2. Hemoglobiny ludzkie- rodzina białek, takich jak mioglobina, spokrewniona z białkami złożonymi (hemoproteinami). Mają budowę tetrameryczną i zawierają dwa łańcuchy α, ale różnią się budową dwóch pozostałych łańcuchów polipeptydowych (łańcuchy 2α, 2x). Struktura drugiego łańcucha polipeptydowego determinuje cechy funkcjonowania tych form Hb. Około 98% hemoglobiny w czerwonych krwinkach dorosłego człowieka to hemoglobina hemoglobina A(łańcuchy 2α, 2p).
Podczas rozwoju płodu działają dwa główne typy hemoglobin: embrionalna Hb(2α, 2ε), który występuje we wczesnych stadiach rozwoju płodu, oraz hemoglobina F (płód)- (2α, 2γ), która zastępuje wczesną hemoglobinę płodową w szóstym miesiącu rozwoju wewnątrzmacicznego i dopiero po urodzeniu zastępuje ją Hb A.
HB A to białko spokrewnione z mioglobiną (MB) występujące w czerwonych krwinkach dorosłego człowieka. Struktura jej poszczególnych protomerów jest podobna do struktury mioglobiny. Struktury drugorzędowe i trzeciorzędowe protomerów mioglobiny i hemoglobiny są bardzo podobne, mimo że w strukturze pierwszorzędowej ich łańcuchów polipeptydowych identyczne są jedynie 24 reszty aminokwasowe (drugorzędowa struktura protomerów hemoglobiny, podobnie jak mioglobiny, zawiera osiem α-helis, oznaczone literami łacińskimi od A do H, a struktura trzeciorzędowa ma postać zwartej globuli). Jednak w przeciwieństwie do mioglobiny hemoglobina ma strukturę oligomeryczną, składającą się z czterech łańcuchów polipeptydowych połączonych wiązaniami niekowalencyjnymi (ryc. 1.22).
Każdy protomer Hb jest powiązany z częścią niebiałkową – hemem i sąsiednimi protomerami. Połączenie części białkowej Hb z hemem jest podobne do połączenia mioglobiny: w centrum aktywnym białka hydrofobowe części hemu są otoczone hydrofobowymi rodnikami aminokwasowymi, z wyjątkiem His F 8 i His E 7, które znajdują się po obu stronach płaszczyzny hemu i odgrywają podobną rolę w funkcjonowaniu białka i jego wiązaniu z tlenem (patrz budowa mioglobiny).
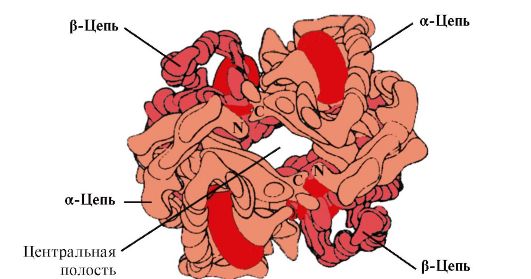 Ryż. 1,22. Oligomeryczna struktura hemoglobiny
Ryż. 1,22. Oligomeryczna struktura hemoglobiny
Oprócz, GisE 7 pełni ważne dodatkowa rola w funkcjonowaniu Nv. Wolny hem ma 25 000 razy większe powinowactwo do CO niż do O2. CO powstaje w organizmie w małych ilościach i ze względu na jego duże powinowactwo do hemu może zakłócać transport O 2 niezbędny do życia komórek. Jednak w składzie hemoglobiny powinowactwo hemu do tlenku węgla przekracza powinowactwo do O 2 tylko 200 razy ze względu na obecność His E 7 w centrum aktywnym. Pozostała część tego aminokwasu stwarza optymalne warunki wiązania hemu z O 2 i osłabia oddziaływanie hemu z CO.
5. Główną funkcją HB jest transport O2 z płuc do tkanek. W przeciwieństwie do monomerycznej mioglobiny, która ma bardzo duże powinowactwo do O2 i pełni funkcję magazynowania tlenu w czerwonych mięśniach, oligomeryczna struktura hemoglobiny zapewnia:
1) szybkie nasycenie HB tlenem w płucach;
2) zdolność HB do uwalniania tlenu w tkankach przy stosunkowo wysokim ciśnieniu cząstkowym O 2 (20–40 mm Hg);
3) możliwość regulacji powinowactwa Hb do O 2.
6. Wspólne zmiany w konformacji protomerów hemoglobiny przyspieszają wiązanie O 2 w płucach i jego uwalnianie do tkanek. W płucach wysokie ciśnienie parcjalne O2 sprzyja jego wiązaniu z Hb w miejscu aktywnym czterech protomerów (2α i 2β). Aktywne centrum każdego protomeru, podobnie jak w mioglobinie, znajduje się pomiędzy dwiema α-helisami (F i E) w kieszeni hydrofobowej. Zawiera część niebiałkową - hem, połączoną z częścią białkową wieloma słabymi oddziaływaniami hydrofobowymi i jednym silnym wiązaniem pomiędzy hemem Fe 2 + i His F 8 (patrz ryc. 1.21).
W deoksyhemoglobinie, dzięki temu wiązaniu z His F 8, atom Fe 2 + wystaje z płaszczyzny hemu w kierunku histydyny. Wiązanie O 2 z Fe 2 + zachodzi po drugiej stronie hemu w regionie His E 7 przy użyciu pojedynczego wolnego wiązania koordynacyjnego. Jego E 7 zapewnia optymalne warunki wiązania O 2 z żelazem hemowym.
Dodatek O2 do atomu Fe+2 jednego protomeru powoduje jego ruch do płaszczyzny hemu, po czym następuje związana z nim reszta histydyny
 Ryż. 1,23. Zmiana konformacji protomeru hemoglobiny w połączeniu z O2
Ryż. 1,23. Zmiana konformacji protomeru hemoglobiny w połączeniu z O2
Prowadzi to do zmiany konformacji wszystkich łańcuchów polipeptydowych ze względu na ich labilność konformacyjną. Zmiana konformacji pozostałych łańcuchów ułatwia ich interakcję z kolejnymi cząsteczkami O 2 .
Czwarta cząsteczka O 2 przyłącza się do hemoglobiny 300 razy łatwiej niż pierwsza (ryc. 1.24).
 Ryż. 1,24. Spółdzielcze zmiany w konformacji protomerów hemoglobiny podczas jej interakcji z O2
Ryż. 1,24. Spółdzielcze zmiany w konformacji protomerów hemoglobiny podczas jej interakcji z O2
W tkankach każda kolejna cząsteczka O 2 jest odcinana łatwiej niż poprzednia, także na skutek kooperatywnych zmian w konformacji protomerów.
7. CO 2 i H+ powstające podczas katabolizmu substancji organicznych zmniejszają powinowactwo hemoglobiny do O 2 proporcjonalnie do ich stężenia. Energia niezbędna do funkcjonowania komórek wytwarzana jest przede wszystkim w mitochondriach podczas utleniania substancji organicznych przy użyciu O 2 dostarczanego z płuc przez hemoglobinę. W wyniku utleniania substancji organicznych powstają końcowe produkty ich rozkładu: CO 2 i K 2 O, których ilość jest proporcjonalna do intensywności zachodzących procesów utleniania.
CO 2 dyfunduje z komórek do krwi i przenika do krwinek czerwonych, gdzie pod wpływem enzymu karbanhydrazy przekształca się w kwas węglowy. Ten słaby kwas dysocjuje na proton i jon wodorowęglanowy.
H+ są w stanie przyłączyć się do Jego rodników 14 6 w łańcuchach α i β hemoglobiny, tj. w obszarach odległych od hemu. Protonowanie hemoglobiny zmniejsza jej powinowactwo do O 2, sprzyja usuwaniu O 2 z oksyHb, tworzeniu deoksyHb i zwiększa dopływ tlenu do tkanek proporcjonalnie do liczby utworzonych protonów (ryc. 1.25).
Wzrost ilości uwalnianego tlenu w zależności od wzrostu stężenia H+ w czerwonych krwinkach nazywany jest efektem Bohra (nazwanym na cześć duńskiego fizjologa Christiana Bohra, który jako pierwszy odkrył ten efekt).
W płucach wysokie ciśnienie parcjalne tlenu sprzyja jego wiązaniu z deoksyHb, co zmniejsza powinowactwo białka do H +. Uwolnione protony pod działaniem kwasu węglowego reagują z wodorowęglanami, tworząc CO 2 i H 2 O
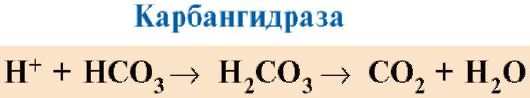
 Ryż. 1,25. Zależność powinowactwa Hb do O 2 od stężenia CO 2 i protonów (efekt Bohra):
Ryż. 1,25. Zależność powinowactwa Hb do O 2 od stężenia CO 2 i protonów (efekt Bohra):
A- wpływ stężenia CO 2 i H+ na uwalnianie O 2 z kompleksu z HB (efekt Bohra); B- natlenienie deoksyhemoglobiny w płucach, tworzenie i uwalnianie CO2.
Powstały CO 2 przedostaje się do przestrzeni pęcherzykowej i jest usuwany wraz z wydychanym powietrzem. Zatem ilość tlenu uwalnianego przez hemoglobinę w tkankach jest regulowana przez produkty katabolizmu substancji organicznych: im intensywniejszy rozkład substancji, na przykład podczas wysiłku fizycznego, tym wyższe stężenie CO 2 i H + oraz tym więcej tlenu tkanki otrzymują w wyniku zmniejszenia powinowactwa Hb do O2.
8. Allosteryczna regulacja powinowactwa Hb do O2 przez ligand – 2,3-bisfosfoglicerynian. W erytrocytach allosteryczny ligand hemoglobiny, 2,3-bisfosfoglicerynian (2,3-BPG), jest syntetyzowany z produktu utleniania glukozy - 1,3-bisfosfoglicerynianu. W normalnych warunkach stężenie 2,3-BPG jest wysokie i porównywalne ze stężeniem Hb. 2,3-BPG ma silny ładunek ujemny -5.
 Bisfosfoglicerynian w naczyniach włosowatych tkanek, wiążąc się z deoksyhemoglobiną, zwiększa uwalnianie tlenu do tkanek, zmniejszając powinowactwo Hb do O2.
Bisfosfoglicerynian w naczyniach włosowatych tkanek, wiążąc się z deoksyhemoglobiną, zwiększa uwalnianie tlenu do tkanek, zmniejszając powinowactwo Hb do O2.
W środku tetramerycznej cząsteczki hemoglobiny znajduje się wnęka. Tworzą go reszty aminokwasowe wszystkich czterech protomerów (patrz ryc. 1.22). W naczyniach włosowatych tkanek protonowanie Hb (efekt Bohra) prowadzi do zerwania wiązania pomiędzy żelazem hemowym i O2. W cząsteczce
deoksyhemoglobina w porównaniu do oksyhemoglobiny pojawiają się dodatkowe wiązania jonowe łączące protomery, w wyniku czego zwiększają się wymiary jamy centralnej w porównaniu z oksyhemoglobiną. Jama centralna jest miejscem przyłączania 2,3-BPG do hemoglobiny. Ze względu na różnicę w wielkości jamy centralnej, 2,3-BPG może przyłączać się jedynie do deoksyhemoglobiny.
2,3-BPG oddziałuje z hemoglobiną w miejscu odległym od aktywnych centrów białka i do którego należy allosteryczny(regulacyjne) ligandy i centralna wnęka Hb centrum allosteryczne. 2,3-BPG ma silny ładunek ujemny i oddziałuje z pięcioma dodatnio naładowanymi grupami dwóch łańcuchów β Hb: N-końcową grupą α-aminową Val i rodnikami Lys 82 His 143 (ryc. 1.26).
 Ryż. 1,26. BPG w centralnej jamie deoksyhemoglobiny
Ryż. 1,26. BPG w centralnej jamie deoksyhemoglobiny
BPG wiąże się z trzema dodatnio naładowanymi grupami na każdej nici β.
W naczyniach włosowatych powstająca deoksyhemoglobina oddziałuje z 2,3-BPG i pomiędzy dodatnio naładowanymi rodnikami łańcuchów β a ujemnie naładowanymi ligandami tworzą się wiązania jonowe, które zmieniają konformację białka i zmniejszają powinowactwo Hb do O2 . Zmniejszenie powinowactwa Hb do O 2 przyczynia się do bardziej wydajnego uwalniania O 2 do tkanki.
W płucach, przy wysokim ciśnieniu parcjalnym, tlen oddziałuje z Hb, łącząc żelazo hemowe; w tym przypadku zmienia się konformacja białka, zmniejsza się wnęka centralna i 2,3-BPG zostaje wyparty z centrum allosterycznego
Zatem białka oligomeryczne mają nowe właściwości w porównaniu do białek monomerycznych. Przyłączanie ligandów w miejscach
odległe od siebie przestrzennie (allosteryczne), mogą powodować zmiany konformacyjne w całej cząsteczce białka. W wyniku interakcji z ligandami regulatorowymi następuje zmiana konformacji i adaptacja funkcji cząsteczki białka do zmian środowiskowych.
TEMAT 1.5. UTRZYMANIE NAtywnej KONFORMACJI BIAŁEK W WARUNKACH KOMÓRKOWYCH
W komórkach podczas syntezy łańcuchów polipeptydowych, ich transportu przez błony do odpowiednich części komórki, podczas procesu fałdowania (tworzenia konformacji natywnej) i podczas składania białek oligomerycznych, a także podczas ich funkcjonowania, pośrednie W strukturze białka powstają podatne na agregację, niestabilne konformacje. Rodniki hydrofobowe, zwykle ukryte wewnątrz cząsteczki białka w konformacji natywnej, pojawiają się na powierzchni w konformacji niestabilnej i mają tendencję do łączenia się z grupami innych białek, które są słabo rozpuszczalne w wodzie. W komórkach wszystkich znane organizmy Odkryto specjalne białka, które zapewniają optymalne zwijanie białek komórkowych, stabilizują ich natywną konformację podczas funkcjonowania i, co najważniejsze, utrzymują strukturę i funkcje białek wewnątrzkomórkowych w przypadku zaburzeń homeostazy. Białka te nazywane są „opiekunowie” co po francusku oznacza „nianię”.
1. Chaperony molekularne i ich rola w zapobieganiu denaturacji białek.
Chaperony (CH) są klasyfikowane według masy ich podjednostek. Chaperony o dużej masie cząsteczkowej mają masę od 60 do 110 kDa. Wśród nich najczęściej badane są trzy klasy: Sh-60, Sh-70 i Sh-90. Każda klasa obejmuje rodzinę powiązanych białek. Zatem Sh-70 obejmuje białka o masie cząsteczkowej od 66 do 78 kDa. Chaperony o niskiej masie cząsteczkowej mają masę cząsteczkową od 40 do 15 kDa.
Wśród opiekunów są składowy białka, których wysoka podstawowa synteza nie zależy od wpływu stresu na komórki organizmu, oraz indukowalny, którego synteza w normalnych warunkach jest słaba, ale gwałtownie wzrasta pod wpływem stresu. Indukowalne białka opiekuńcze nazywane są także „białkami szoku cieplnego”, ponieważ po raz pierwszy odkryto je w komórkach narażonych na działanie wysokich temperatur. W komórkach, ze względu na duże stężenie białek, spontaniczna reaktywacja częściowo zdenaturowanych białek jest utrudniona. Sh-70 może zapobiegać denaturacji i pomagać w przywracaniu natywnej konformacji białek. Chaperony molekularne-70- wysoce konserwatywna klasa białek występująca we wszystkich częściach komórki: cytoplazmie, jądrze, retikulum endoplazmatycznym, mitochondriach. Na końcu karboksylowym pojedynczego łańcucha polipeptydowego Ш-70 znajduje się region będący rowkiem zdolnym do interakcji z peptydami o długości
od 7 do 9 reszt aminokwasowych wzbogaconych rodnikami hydrofobowymi. Takie regiony w białkach globularnych występują w przybliżeniu co 16 aminokwasów. Sh-70 jest w stanie chronić białka przed inaktywacją temperaturową oraz przywracać konformację i aktywność częściowo zdenaturowanych białek.
2. Rola białek opiekuńczych w zwijaniu białek. Podczas syntezy białka na rybosomie, N-końcowy region polipeptydu jest syntetyzowany przed C-końcowym. Aby utworzyć natywną konformację, wymagana jest pełna sekwencja aminokwasów białka. W procesie syntezy białek chaperony-70, dzięki strukturze swojego centrum aktywnego, są w stanie zamykać obszary polipeptydu podatne na agregację, wzbogacone w hydrofobowe rodniki aminokwasowe, aż do zakończenia syntezy (Rysunek 1.27, A ).
 Ryż. 1,27. Udział białek opiekuńczych w fałdowaniu białek
Ryż. 1,27. Udział białek opiekuńczych w fałdowaniu białek
A - udział chaperonów-70 w zapobieganiu oddziaływaniom hydrofobowym pomiędzy odcinkami syntetyzowanego polipeptydu; B - tworzenie natywnej konformacji białka w kompleksie chaperonowym
Wiele białek wielkocząsteczkowych o złożonej konformacji, takiej jak struktura domeny, składa się w specjalnej przestrzeni utworzonej przez Sh-60. Ř-60 funkcjonują jako kompleks oligomeryczny składający się z 14 podjednostek. Tworzą dwa puste pierścienie, z których każdy składa się z siedmiu podjednostek, pierścienie te są ze sobą połączone. Każda podjednostka Sh-60 składa się z trzech domen: wierzchołkowej (wierzchołkowej), wzbogaconej rodnikami hydrofobowymi skierowanymi w stronę wnęki pierścienia, pośredniej i równikowej (ryc. 1.28).
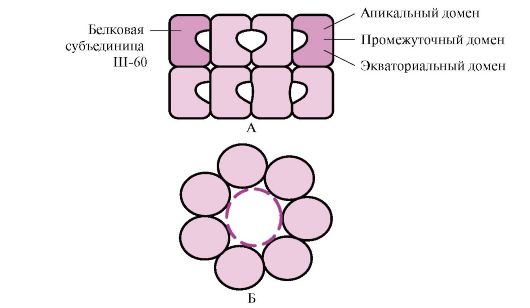 Ryż. 1,28. Struktura kompleksu opiekuńczego składającego się z 14 Ш-60
Ryż. 1,28. Struktura kompleksu opiekuńczego składającego się z 14 Ш-60
A - widok z boku; B - widok z góry
Zsyntetyzowane białka, które mają na powierzchni elementy charakterystyczne dla rozwiniętych cząsteczek, w szczególności rodniki hydrofobowe, dostają się do wnęki pierścieni opiekuńczych. W specyficznym środowisku tych wnęk poszukuje się możliwych konformacji, aż do znalezienia jedynej, najkorzystniejszej energetycznie (rys. 1.27, B). Tworzeniu konformacji i uwalnianiu białek towarzyszy hydroliza ATP w obszarze równikowym. Zazwyczaj takie fałdowanie zależne od opiekunów wymaga znacznej ilości energii.
Oprócz udziału w tworzeniu trójwymiarowej struktury białek i renatywacji częściowo zdenaturowanych białek, chaperony są również niezbędne do zachodzenia tak podstawowych procesów jak składanie białek oligomerycznych, rozpoznawanie i transport zdenaturowanych białek do lizosomów, transport białek przez błony i udział w regulacji aktywności kompleksów białkowych.
TEMAT 1.6. RÓŻNORODNOŚĆ BIAŁEK. RODZINY BIAŁEK: PRZYKŁAD IMMUNOGLOBULIN
1. Białka odgrywają decydującą rolę w życiu poszczególnych komórek i całego organizmu wielokomórkowego, a ich funkcje są zaskakująco różnorodne. Decydują o tym cechy struktury pierwszorzędowej i konformacji białek, unikalna struktura centrum aktywnego oraz zdolność wiązania określonych ligandów.
Tylko bardzo mała część wszystkich możliwych wariantów łańcuchów peptydowych może przyjąć stabilną strukturę przestrzenną; większość
z nich może przyjmować wiele konformacji o w przybliżeniu tej samej energii Gibbsa, ale o różnych właściwościach. Struktura pierwotna większości znanych białek wyselekcjonowana na drodze ewolucji biologicznej zapewnia wyjątkową stabilność jednej z konformacji, która determinuje charakterystykę funkcjonowania tego białka.
2. Rodziny białek. W ciągu jednego gatunki biologiczne podstawienia reszt aminokwasowych mogą prowadzić do pojawienia się różnych białek, które spełniają powiązane funkcje i mają homologiczne sekwencje aminokwasów. Takie pokrewne białka mają uderzająco podobne konformacje: liczbę i względne położenie α-helis i/lub struktur β, a większość zwojów i zagięć łańcuchów polipeptydowych jest podobna lub identyczna. Białka z homologicznymi regionami łańcucha polipeptydowego, podobną konformacją i powiązanymi funkcjami dzieli się na rodziny białek. Przykłady rodzin białek: proteinazy serynowe, rodzina immunoglobulin, rodzina mioglobin.
Proteinazy serynowe- rodzina białek pełniących funkcję enzymów proteolitycznych. Należą do nich enzymy trawienne – chymotrypsyna, trypsyna, elastaza i wiele czynników krzepnięcia krwi. Białka te mają identyczne aminokwasy w 40% swoich pozycji i bardzo podobną konformację (ryc. 1.29).

Ryż. 1,29. Struktury przestrzenne elastazy (A) i chymotrypsyny (B)
Niektóre podstawienia aminokwasów doprowadziły do zmian w specyficzności substratowej tych białek i pojawienia się różnorodności funkcjonalnej w obrębie rodziny.
3. Rodzina immunoglobulin. W funkcjonowaniu układu odpornościowego ogromną rolę odgrywają białka z nadrodziny immunoglobulin, do której zaliczają się trzy rodziny białek:
Przeciwciała (immunoglobuliny);
Receptory limfocytów T;
Białka głównego kompleksu zgodności tkankowej - klasy MHC 1 i 2 (Major Histocompatibility Complex).
Wszystkie te białka mają budowę domenową, składają się z homologicznych domen immunopodobnych i pełnią podobne funkcje: oddziałują z obcymi strukturami, albo rozpuszczonymi we krwi, limfie, płynie międzykomórkowym (przeciwciała), albo znajdującymi się na powierzchni komórek (własnych lub zagraniczny).
4. Przeciwciała- specyficzne białka wytwarzane przez limfocyty B w odpowiedzi na przedostanie się obcej struktury do organizmu, tzw antygen.
Cechy struktury przeciwciał
Najprostsze cząsteczki przeciwciał składają się z czterech łańcuchów polipeptydowych: dwóch identycznych lekkich - L, zawierających około 220 aminokwasów i dwóch identycznych ciężkich - H, składających się z 440-700 aminokwasów. Wszystkie cztery łańcuchy w cząsteczce przeciwciała są połączone wieloma wiązaniami niekowalencyjnymi i czterema wiązaniami dwusiarczkowymi (ryc. 1.30).
Łańcuchy lekkie przeciwciał składają się z dwóch domen: domeny zmiennej (VL), zlokalizowanej w regionie N-końcowym łańcucha polipeptydowego i domeny stałej (CL), zlokalizowanej na C-końcu. Łańcuchy ciężkie mają zwykle cztery domeny: jedną zmienną (VH), zlokalizowaną na N-końcu i trzy domeny stałe (CH1, CH2, CH3) (patrz ryc. 1.30). Każda domena immunoglobulinowa ma nadbudowę arkusza β, w której dwie reszty cysteinowe są połączone wiązaniem dwusiarczkowym.
Pomiędzy dwiema domenami stałymi CH1 i CH2 znajduje się region zawierający dużą liczbę reszt proliny, które zapobiegają tworzeniu się struktury drugorzędowej i oddziaływaniu sąsiadujących łańcuchów H w tym segmencie. Ten region zawiasowy zapewnia elastyczność cząsteczki przeciwciała. Pomiędzy domenami zmiennymi łańcucha ciężkiego i lekkiego znajdują się dwa identyczne miejsca wiązania antygenu (miejsca aktywne wiązania antygenów), dlatego takie przeciwciała często nazywane są biwalenty. W wiązaniu antygenu z przeciwciałem nie bierze udziału cała sekwencja aminokwasów regionów zmiennych obu łańcuchów, ale jedynie 20-30 aminokwasów znajdujących się w regionach hiperzmiennych każdego łańcucha. To właśnie te regiony określają unikalną zdolność każdego typu przeciwciał do interakcji z odpowiednim antygenem komplementarnym.
Przeciwciała są jedną z linii obrony organizmu przed inwazją obcych organizmów. Ich funkcjonowanie można podzielić na dwa etapy: pierwszy etap to rozpoznanie i związanie antygenu na powierzchni obcych organizmów, co jest możliwe dzięki obecności w strukturze przeciwciała miejsc wiążących antygen; drugi etap to inicjacja procesu inaktywacji i zniszczenia antygenu. Specyficzność drugiego etapu zależy od klasy przeciwciał. Wyróżnia się pięć klas łańcuchów ciężkich, różniących się między sobą budową domen stałych: α, δ, ε, γ i μ, według których wyróżnia się pięć klas immunoglobulin: A, D, E, G i M.
Cechy strukturalne łańcuchów ciężkich nadają regionom zawiasowym i regionom C-końcowym łańcuchów ciężkich konformację charakterystyczną dla każdej klasy. Po związaniu antygenu z przeciwciałem zmiany konformacyjne w domenach stałych określają ścieżkę usuwania antygenu.
 Ryż. 1. 30. Struktura domenowa IgG
Ryż. 1. 30. Struktura domenowa IgG
Immunoglobuliny M
Immunoglobuliny M mają dwie formy.
Forma monomeryczna- I klasa przeciwciał wytwarzanych przez rozwijające się limfocyty B. Następnie wiele limfocytów B zaczyna wytwarzać przeciwciała innej klasy, ale z tym samym miejscem wiązania antygenu. IgM jest osadzone w błonie i działa jako receptor rozpoznający antygen. Integracja IgM z błoną komórkową jest możliwa dzięki obecności 25 hydrofobowych reszt aminokwasowych w części ogonowej regionu.
Wydzielnicza postać IgM zawiera pięć monomerycznych podjednostek połączonych ze sobą wiązaniami dwusiarczkowymi i dodatkowym polipeptydowym łańcuchem J (ryc. 1.31). Łańcuchy ciężkie monomerów tej postaci nie zawierają ogona hydrofobowego. Pentamer ma 10 miejsc wiązania antygenu i dlatego skutecznie rozpoznaje i usuwa antygen, który jako pierwszy dostanie się do organizmu. Wydzielnicza postać IgM to główna klasa przeciwciał wydzielanych do krwi podczas pierwotnej odpowiedzi immunologicznej. Wiązanie IgM z antygenem zmienia konformację IgM i indukuje jej związanie z pierwszym białkowym składnikiem układu dopełniacza (układ dopełniacza to zespół białek biorących udział w niszczeniu antygenu) i aktywację tego układu. Jeżeli antygen znajduje się na powierzchni drobnoustroju, układ dopełniacza powoduje przerwanie integralności błony komórkowej i śmierć komórki bakteryjnej.
Immunoglobuliny G
Ilościowo ta klasa immunoglobulin przeważa we krwi (75% wszystkich Ig). IgG – monomery, główna klasa przeciwciał wydzielanych do krwi podczas wtórnej odpowiedzi immunologicznej. Po interakcji IgG z antygenami powierzchniowymi mikroorganizmów kompleks antygen-przeciwciało jest w stanie wiązać i aktywować białka układu dopełniacza lub oddziaływać ze specyficznymi receptorami makrofagów i neutrofili. Interakcja z fagocytami prowadzi
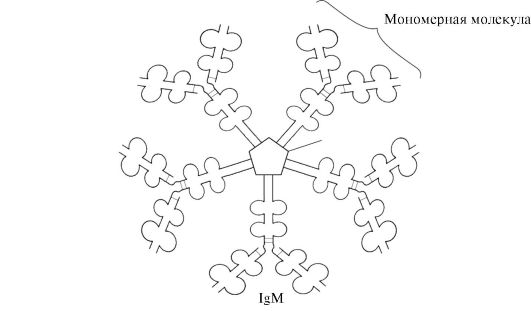 Ryż. 1.31. Struktura wydzielniczej formy IgM
Ryż. 1.31. Struktura wydzielniczej formy IgM
na absorpcję kompleksów antygen-przeciwciało i ich zniszczenie w fagosomach komórkowych. IgG to jedyna klasa przeciwciał, która jest w stanie przeniknąć przez barierę łożyskową i zapewnić wewnątrzmaciczną ochronę płodu przed infekcjami.
Immunoglobuliny A
Główna klasa przeciwciał występujących w wydzielinach (mleko, ślina, wydzieliny dróg oddechowych i jelit). IgA jest wydzielana głównie w postaci dimerycznej, gdzie monomery są połączone ze sobą dodatkowym łańcuchem J (ryc. 1.32).
IgA nie oddziałuje z układem dopełniacza i komórkami fagocytarnymi, ale wiążąc się z mikroorganizmami, przeciwciała uniemożliwiają ich przyłączenie do komórek nabłonkowych i przenikanie do organizmu.
Immunoglobuliny E
Immunoglobuliny E są reprezentowane przez monomery, w których ciężkie łańcuchy ε zawierają, podobnie jak łańcuchy μ immunoglobulin M, jedną domenę zmienną i cztery stałe. Po wydzieleniu IgE wiąże się ze swoimi
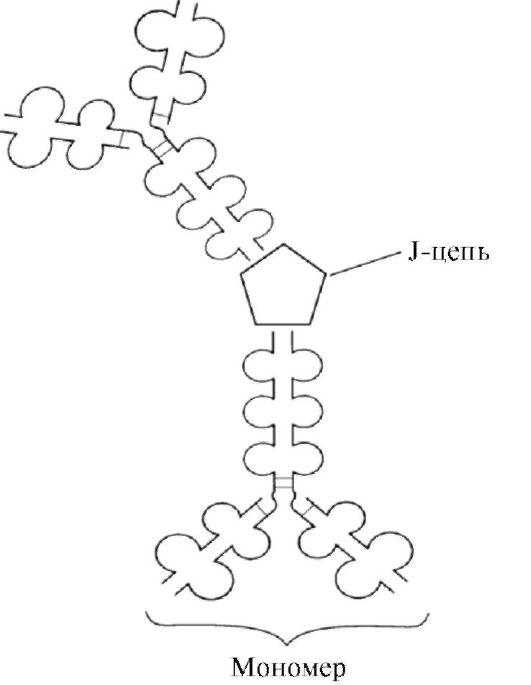 Ryż. 1,32. Struktura IgA
Ryż. 1,32. Struktura IgA
Regiony C-końcowe z odpowiednimi receptorami na powierzchni komórek tucznych i bazofilów. W rezultacie stają się receptorami dla antygenów na powierzchni tych komórek (ryc. 1.33).
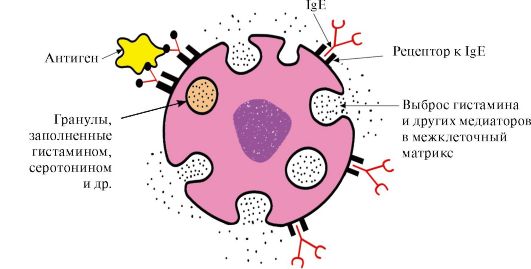 Ryż. 1,33. Oddziaływanie IgE z antygenem na powierzchni komórek tucznych
Ryż. 1,33. Oddziaływanie IgE z antygenem na powierzchni komórek tucznych
Po przyłączeniu się antygenu do odpowiednich miejsc wiązania antygenu IgE, komórki otrzymują sygnał do wydzielania substancji biologicznie czynnych (histamina, serotonina), które w dużej mierze odpowiadają za rozwój reakcji zapalnej i manifestację reakcji alergicznych, takich jak np. astma, pokrzywka, katar sienny.
Immunoglobuliny D
Immunoglobuliny D występują w bardzo małych ilościach w surowicy i są monomerami. Ciężkie łańcuchy δ mają jedną domenę zmienną i trzy domeny stałe. IgD działają jako receptory dla limfocytów B; inne funkcje są nadal nieznane. Oddziaływanie specyficznych antygenów z receptorami na powierzchni limfocytów B (IgD) prowadzi do przekazania tych sygnałów do komórki i aktywacji mechanizmów zapewniających proliferację danego klonu limfocytów.
TEMAT 1.7. WŁAŚCIWOŚCI FIZYCZNE I CHEMICZNE BIAŁEK ORAZ METODY ICH SEPARACJI
1. Poszczególne białka różnią się właściwościami fizycznymi i chemicznymi:
Kształt cząsteczek;
Waga molekularna;
Całkowity ładunek, którego wartość zależy od stosunku grup anionowych i kationowych aminokwasów;
Stosunek polarnych i niepolarnych rodników aminokwasowych na powierzchni cząsteczek;
Stopnie odporności na różne czynniki denaturujące.
2. Rozpuszczalność białka zależy od właściwości wymienionych białek, a także od składu środowiska, w którym białko jest rozpuszczane (wartości pH, skład soli, temperatura, obecność innych substancji organicznych mogących oddziaływać z białkiem). Wielkość ładunku cząsteczek białek jest jednym z czynników wpływających na ich rozpuszczalność. Kiedy ładunek w punkcie izoelektrycznym zostanie utracony, białka łatwiej agregują i wytrącają się. Jest to szczególnie typowe dla białek zdenaturowanych, w których na powierzchni pojawiają się hydrofobowe rodniki aminokwasowe.
Na powierzchni cząsteczki białka znajdują się zarówno dodatnio, jak i ujemnie naładowane rodniki aminokwasów. Liczba tych grup, a co za tym idzie całkowity ładunek białek, zależy od pH ośrodka, tj. stosunek stężeń grup H+ - i OH -. W kwaśnym środowisku Wzrost stężenia H+ prowadzi do zahamowania dysocjacji grup karboksylowych -COO - + H+ > - COOH i zmniejszenia ładunku ujemnego białek. W środowisku zasadowym wiązanie nadmiaru OH - przez protony powstałe podczas dysocjacji grup aminowych -NH 3 + + OH - - NH 2 + H 2 O z utworzeniem wody prowadzi do zmniejszenia ładunku dodatniego białek . Nazywa się wartość pH, przy której białko ma ładunek zerowy netto punkt izoelektryczny (IEP). W IET liczba grup naładowanych dodatnio i ujemnie jest taka sama, tj. białko jest w stanie izoelektrycznym.
3. Rozdzielanie poszczególnych białek. Cechy budowy i funkcjonowania organizmu zależą od zestawu syntetyzowanych w nim białek. Badanie struktury i właściwości białek nie jest możliwe bez wyizolowania ich z komórki i oczyszczenia z innych białek i cząsteczek organicznych. Etapy izolacji i oczyszczania poszczególnych białek:
Zniszczenie komórek badanej tkanki i uzyskania homogenatu.
Podział homogenatu na frakcje przez wirowanie, uzyskując frakcję jądrową, mitochondrialną, cytozolową lub inną zawierającą pożądane białko.
Selektywna denaturacja termiczna- krótkotrwałe ogrzewanie roztworu białka, podczas którego można usunąć część zdenaturowanych zanieczyszczeń białkowych (o ile białko jest stosunkowo termostabilne).
Solenie. Różne białka wytrącają się przy różnym stężeniu soli w roztworze. Stopniowo zwiększając stężenie soli, można otrzymać szereg odrębnych frakcji, w których w jednej z nich przeważa zawartość izolowanego białka. Do frakcjonowania białek najczęściej stosuje się siarczan amonu. Białka o najmniejszej rozpuszczalności wytrącają się przy niskich stężeniach soli.
Filtracja żelowa- metoda przesiewania cząsteczek przez spęczniałe granulki Sephadexu (trójwymiarowe łańcuchy polisacharydowe dekstranu posiadające pory). Szybkość, z jaką białka przechodzą przez kolumnę wypełnioną Sephadexem, będzie zależała od ich masy cząsteczkowej: im mniejsza masa cząsteczek białka, tym łatwiej wnikają one do granulek i dłużej tam pozostają; im większa masa, tym szybciej wymywają się z kolumna.
Ultrawirowanie- metoda polegająca na umieszczeniu białek w probówce wirówkowej w rotorze ultrawirówki. Kiedy rotor się obraca, szybkość sedymentacji białek jest proporcjonalna do ich masy cząsteczkowej: frakcje białek cięższych znajdują się bliżej dna probówki, lżejsze - bliżej powierzchni.
Elektroforeza- metoda oparta na różnicach prędkości ruchu białek w polu elektrycznym. Wartość ta jest proporcjonalna do ładunku białek. Elektroforezę białek przeprowadza się na papierze (w tym przypadku prędkość ruchu białek jest proporcjonalna tylko do ich ładunku) lub w żelu poliakryloamidowym o określonej wielkości porów (szybkość ruchu białek jest proporcjonalna do ich ładunku i masy cząsteczkowej) .
Chromatografia jonowymienna- metoda frakcjonowania polegająca na wiązaniu zjonizowanych grup białek z przeciwnie naładowanymi grupami żywic jonowymiennych (nierozpuszczalnych materiałów polimerowych). Siła wiązania białka z żywicą jest proporcjonalna do ładunku białka. Białka zaadsorbowane na polimerze jonowymiennym można wypłukać wraz ze wzrostem stężenia roztworów NaCl; im niższy ładunek białka, tym niższe stężenie NaCl wymagane do wypłukania białka związanego z grupami jonowymi żywicy.
Chromatografii powinowactwa- najbardziej specyficzna metoda izolacji poszczególnych białek.Ligand białka jest kowalencyjnie przyłączony do obojętnego polimeru. Kiedy roztwór białka przepuszczany jest przez kolumnę z polimerem, na kolumnie adsorbowane jest wyłącznie białko specyficzne dla danego liganda, ze względu na komplementarne wiązanie białka z ligandem.
Dializa- metoda służąca do usuwania związków drobnocząsteczkowych z roztworu izolowanego białka. Metoda opiera się na niezdolności białek do przejścia przez półprzepuszczalną membranę, w przeciwieństwie do substancji o niskiej masie cząsteczkowej. Służy do oczyszczania białek z zanieczyszczeń drobnocząsteczkowych, np. soli po wysalaniu.
ZADANIA NA PRACĘ POZASZKOLNĄ
1. Wypełnij tabelę. 1.4.
Tabela 1.4. Analiza porównawcza budowy i funkcji powiązanych białek - mioglobiny i hemoglobiny
a) zapamiętaj strukturę aktywnego centrum Mb i Hb. Jaką rolę odgrywają hydrofobowe rodniki aminokwasowe w tworzeniu centrów aktywnych tych białek? Opisać budowę centrum aktywnego Mb i Hb oraz mechanizmy przyłączania do niego ligandów. Jaką rolę odgrywają reszty His F 8 i His E 7 w funkcjonowaniu centrum aktywnego Mv iHv?
b) jakie nowe właściwości w porównaniu z monomeryczną mioglobiną ma blisko spokrewnione białko oligomeryczne, hemoglobina? Wyjaśnić rolę kooperatywnych zmian w konformacji protomerów w cząsteczce hemoglobiny, wpływ stężenia CO 2 i protonów na powinowactwo hemoglobiny do tlenu, a także rolę 2,3-BPG w allosterycznej regulacji funkcji Hb .
2. Scharakteryzuj chaperony molekularne, zwracając uwagę na związek pomiędzy ich strukturą i funkcją.
3. Jakie białka dzielimy na rodziny? Na przykładzie rodziny immunoglobulin zidentyfikuj podobne cechy strukturalne i powiązane funkcje białek tej rodziny.
4. Oczyszczone pojedyncze białka są często wymagane do celów biochemicznych i leczniczych. Wyjaśnij, na jakich właściwościach fizykochemicznych białek opierają się metody stosowane do ich rozdzielania i oczyszczania.
ZADANIA SAMOKONTROLI
1. Wybierz prawidłowe odpowiedzi.
Funkcje hemoglobiny:
A. Transport O 2 z płuc do tkanek B. Transport H + z tkanek do płuc
B. Utrzymanie stałego pH krwi. D. Transport CO 2 z płuc do tkanek
D. Transport CO 2 z tkanek do płuc
2. Wybierz prawidłowe odpowiedzi. Ligandα -protomer Hb to: A. Hem
B. Tlen
B. CO G. 2,3-BPG
D. β-Protomer
3. Wybierz prawidłowe odpowiedzi.
Hemoglobina w przeciwieństwie do mioglobiny:
A. Ma strukturę czwartorzędową
B. Strukturę drugorzędową reprezentują jedynie α-helisy
B. Należy do białek złożonych
D. Oddziałuje z ligandem allosterycznym. D. Kowalencyjnie związany z hemem
4. Wybierz prawidłowe odpowiedzi.
Powinowactwo Hb do O2 maleje:
A. Gdy dodana zostanie jedna cząsteczka O 2. B. Gdy zostanie usunięta jedna cząsteczka O 2
B. Podczas interakcji z 2,3-BPG
D. Po przyłączeniu do protomerów H + D. Gdy stężenie 2,3-BPG maleje
5. Mecz.
Typy HB charakteryzują się:
A. W formie deoksy tworzy agregaty włókniste B. Zawiera dwa łańcuchy α i dwa δ
B. Dominująca forma Hb w erytrocytach dorosłych D. Zawiera hem z Fe+ 3 w centrum aktywnym
D. Zawiera dwa łańcuchy α i dwa łańcuchy γ 1. HbA 2.
6. Mecz.
Ligandy Hb:
A. Wiąże się z Hb w centrum allosterycznym
B. Ma bardzo duże powinowactwo do miejsca aktywnego Hb
B. Łącząc zwiększa powinowactwo Hb do O 2 G. Utlenia Fe+ 2 do Fe+ 3
D. Tworzy wiązanie kowalencyjne z hisF8
7. Wybierz prawidłowe odpowiedzi.
Opiekunowie:
A. Białka obecne we wszystkich częściach komórki
B. Synteza wzrasta pod wpływem stresu
B. Uczestniczyć w hydrolizie zdenaturowanych białek
D. Uczestniczyć w utrzymaniu natywnej konformacji białek
D. Tworzą organelle, w których powstaje konformacja białek.
8. Mecz. Immunoglobuliny:
A. Forma wydzielnicza jest pentameryczna.
B. Klasa Ig, która przenika przez barierę łożyskową
B. Ig - receptor komórek tucznych
D. Główna klasa Ig obecna w wydzielinach komórek nabłonkowych. D. Receptor limfocytów B, którego aktywacja zapewnia proliferację komórek
9. Wybierz prawidłowe odpowiedzi.
Immunoglobuliny E:
A. Wytwarzane przez makrofagi. B. Mają ciężkie łańcuchy ε.
B. Osadzone w błonie limfocytów T
D. Działają jako błonowe receptory antygenów na komórkach tucznych i bazofilach
D. Odpowiedzialny za reakcje alergiczne
10. Wybierz prawidłowe odpowiedzi.
Metoda rozdziału białek opiera się na różnicach w ich masie cząsteczkowej:
A. Filtracja żelowa
B. Ultrawirowanie
B. Elektroforeza w żelu poliakryloamidowym. D. Chromatografia jonowymienna
D. Chromatografia powinowactwa
11. Wybierz poprawną odpowiedź.
Metoda rozdziału białek opiera się na różnicach w ich rozpuszczalności w wodzie:
A. Filtracja żelowa B. Wysalanie
B. Chromatografia jonowymienna. D. Chromatografia powinowactwa
D. Elektroforeza w żelu poliakryloamidowym
STANDARDY ODPOWIEDZI NA „ZADANIA SAMOKONTROLI”
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-B
PODSTAWOWE TERMINY I POJĘCIA
1. Białka oligomeryczne, protomer, czwartorzędowa struktura białek
2. Kooperacyjne zmiany w konformacji protomeru
3. Efekt Bohra
4. Allosteryczna regulacja funkcji białek, centrum allosteryczne i efektor allosteryczny
5. Chaperony molekularne, białka szoku cieplnego
6. Rodziny białek (proteazy serynowe, immunoglobuliny)
7. Zależność IgM-, G-, E-, A-struktura-funkcja
8. Ładunek całkowity białek, punkt izoelektryczny białek
9. Elektroforeza
10. Solenie
11. Filtracja żelowa
12. Chromatografia jonowymienna
13. Ultrawirowanie
14. Chromatografia powinowactwa
15. Elektroforeza białek osocza krwi
ZADANIA DO PRACY W KLASIE
1. Porównaj zależności stopnia nasycenia hemoglobiny (Hb) i mioglobiny (Mb) tlenem od jej ciśnienia parcjalnego w tkankach
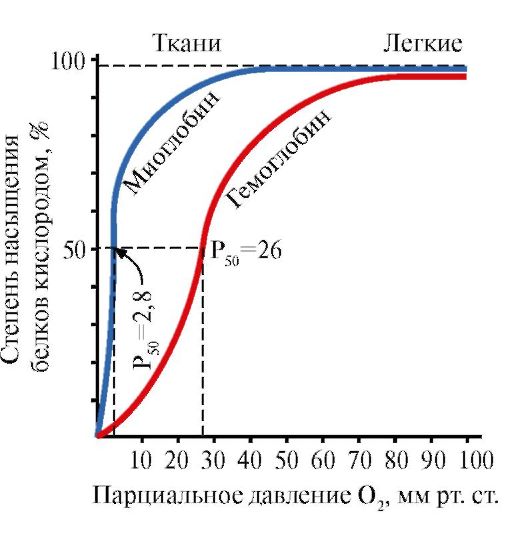 Ryż. 1,34. Zależność nasycenia Mv iNHtlenu z jego ciśnienia cząstkowego
Ryż. 1,34. Zależność nasycenia Mv iNHtlenu z jego ciśnienia cząstkowego
Należy pamiętać, że kształt krzywych nasycenia białka tlenem jest inny: dla mioglobiny - hiperbola, dla hemoglobiny - kształt esowaty.
1. porównać wartości ciśnienia cząstkowego tlenu, przy którym Mb i Hb są nasycone O 2 o 50%. Które z tych białek ma większe powinowactwo do O2?
2. Jakie cechy strukturalne Mb decydują o jego wysokim powinowactwie do O 2?
3. Jakie cechy strukturalne HB pozwalają na uwalnianie O2 w kapilarach spoczynkowych tkanek (przy stosunkowo wysokim ciśnieniu parcjalnym O2) i gwałtownie zwiększają to uwalnianie w pracujących mięśniach? Jaka właściwość białek oligomerycznych zapewnia taki efekt?
4. Oblicz, jaką ilość O 2 (w%) utlenionej hemoglobiny oddaje mięśnie odpoczywające i pracujące?
5. wyciągać wnioski na temat związku pomiędzy strukturą białka a jego funkcją.
2. Ilość tlenu uwalnianego przez hemoglobinę w naczyniach włosowatych zależy od intensywności procesów katabolicznych w tkankach (efekt Bohra). W jaki sposób zmiany w metabolizmie tkanek regulują powinowactwo Hb do O2? Wpływ CO 2 i H+ na powinowactwo Hb do O 2
 1. opisać efekt Bohra.
1. opisać efekt Bohra.
2. w jakim kierunku przebiega proces pokazany na schemacie:
a) w naczyniach włosowatych płuc;
b) w naczyniach włosowatych tkankowych?
3. Jakie jest fizjologiczne znaczenie efektu Bohra?
4. Dlaczego oddziaływanie Hb z H+ w miejscach odległych od hemu zmienia powinowactwo białka do O 2?
3. Powinowactwo Hb do O2 zależy od stężenia jej ligandu – 2,3-bisfosfoglicerynianu, który jest allosterycznym regulatorem powinowactwa Hb do O2. Dlaczego interakcja ligandu w miejscu odległym od miejsca aktywnego wpływa na funkcję białka? W jaki sposób 2,3-BPG reguluje powinowactwo Hb do O2? Aby rozwiązać problem, odpowiedz na następujące pytania:
1. gdzie i z czego syntetyzowany jest 2,3-bisfosfoglicerynian (2,3-BPG)? Napisz jego wzór, wskaż ładunek tej cząsteczki.
2. Z jaką formą hemoglobiny (oksy czy deoksy) wchodzi w interakcję BPG i dlaczego? W jakiej części cząsteczki Hb zachodzi interakcja?
3. w jakim kierunku przebiega proces pokazany na schemacie?
a) w naczyniach włosowatych tkankowych;
b) w naczyniach włosowatych płuc?
4. gdzie stężenie kompleksu powinno być wyższe
Nv-2,3-BFG:
a) w naczyniach włosowatych mięśni w stanie spoczynku,
b) w naczyniach włosowatych pracujących mięśni (pod warunkiem takiego samego stężenia BPG w erytrocytach)?
5. Jak zmieni się powinowactwo HB do tlenu, gdy człowiek przystosuje się do warunków wysokogórskich, jeśli wzrośnie stężenie BPG w erytrocytach? Jakie jest fizjologiczne znaczenie tego zjawiska?
4. Zniszczenie 2,3-BPG podczas przechowywania zakonserwowanej krwi upośledza funkcje HB. Jak zmieni się powinowactwo HB do O 2 w zakonserwowanej krwi, jeśli stężenie 2,3-BPG w erytrocytach może spaść z 8 do 0,5 mmol/l. Czy można przetaczać taką krew ciężko chorym, jeśli stężenie 2,3-BPG zostanie przywrócone dopiero po trzech dniach? Czy możliwe jest przywrócenie funkcji czerwonych krwinek poprzez dodanie do krwi 2,3-BPG?
5. Zapamiętaj strukturę najprostszych cząsteczek immunoglobulin. Jaką rolę odgrywają immunoglobuliny w układzie odpornościowym? Dlaczego Igs często nazywane są biwalentami? Jak struktura Ig wiąże się z ich funkcją? (Opisz na przykładzie klasy immunoglobulin.)
Właściwości fizykochemiczne białek i metody ich rozdzielania.
6. Jak ładunek netto białka wpływa na jego rozpuszczalność?
a) określić całkowity ładunek peptydu przy pH 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis
b) jak zmieni się ładunek tego peptydu przy pH >7, pH<7, рН <<7?
c) jaki jest punkt izoelektryczny białka (IEP) i w jakim środowisku się ono znajduje?
IET tego peptydu?
d) przy jakiej wartości pH będzie zaobserwowana najmniejsza rozpuszczalność tego peptydu.
7. Dlaczego kwaśne mleko, w przeciwieństwie do świeżego mleka, „zsiada się” po ugotowaniu (tj. wytrąca się kazeina białka mleka)? W świeżym mleku cząsteczki kazeiny mają ładunek ujemny.
8. Do separacji poszczególnych białek stosuje się filtrację żelową. Mieszaninę zawierającą białka A, B, C o masach cząsteczkowych odpowiednio 160 000, 80 000 i 60 000 poddano analizie metodą filtracji żelowej (ryc. 1.35). Spęcznione granulki żelu przepuszczają białka o masie cząsteczkowej mniejszej niż 70 000. Na jakiej zasadzie leży ta metoda separacji? Który wykres poprawnie przedstawia wyniki frakcjonowania? Wskaż kolejność, w jakiej białka A, B i C są uwalniane z kolumny.
 Ryż. 1,35. Zastosowanie filtracji żelowej do separacji białek
Ryż. 1,35. Zastosowanie filtracji żelowej do separacji białek
9. Na ryc. 1.36, A przedstawia schemat elektroforezy na papierze białek surowicy krwi osoby zdrowej. Względne ilości frakcji białkowych otrzymanych tą metodą wynoszą: albuminy 54-58%, α1-globuliny 6-7%, α2-globuliny 8-9%, β-globuliny 13%, γ-globuliny 11-12%.
 Ryż. 1.36 Elektroforeza na papierze białek osocza krwi osoby zdrowej (A) i pacjenta (B)
Ryż. 1.36 Elektroforeza na papierze białek osocza krwi osoby zdrowej (A) i pacjenta (B)
I - γ-globuliny; II - β-globuliny; III -α 2 -globulina; IV -α 2 -globulina; V - albuminy
Wielu chorobom towarzyszą ilościowe zmiany w składzie białek surowicy (dyproteinemia). Charakter tych zmian jest brany pod uwagę przy stawianiu diagnozy oraz ocenie ciężkości i stadium choroby.
Korzystając z danych podanych w tabeli. 1.5, zgadnij o chorobie, którą charakteryzuje profil elektroforetyczny przedstawiony na ryc. 1,36.
Tabela 1.5. Zmiany stężenia białek surowicy w patologii









- Eksperymentalne metody rejestracji cząstek elementarnych
- Z czego słynie szwedzki mecz?
- Najlepsze haubice samobieżne rosyjskiej artylerii
- Fiodor Pietrowicz Litke: drugie opłynięcie świata
- Otwarto pierwszą aptekę
- Znakomity podróżnik i naukowiec Fiodor Pietrowicz Litke Znaczek wydany na cześć wyprawy Litke
- Unia Zakonu Kawalerów Mieczowych z Zakonem Krzyżackim
- Ciasto na pizzę – sekrety kuchni włoskiej
- Jak zrobić koszyczki z ciasta kruchego
- Mięso warstwami ze smalcem w łuskach cebuli – wyjdzie, za uszy nie da się go oderwać. Smalec gotowany z warstwą
- Sos wieprzowy - przepisy na sos wieprzowy na patelni i w powolnej kuchence
- Jak zapłacić podatek transportowy dla osób prawnych
- Karta do indywidualnego rozliczania kwot naliczonych wpłat i pozostałych wynagrodzeń oraz kwot naliczonych składek ubezpieczeniowych Karta rocznych składek ubezpieczeniowych
- Błąd podczas wypełniania konturu
- Audyt biurka: zmiany
- Warunki przeprowadzenia eksperymentu rufowego
- Stan systemu i procesy
- Rozwój działalności badawczej studentów Aleksieja Siergiejewicza Obuchowa
- Mars 4. planeta Układu Słonecznego
- Układy narządów człowieka