Budowa i funkcje błon komórkowych. Budowa i funkcje błon biologicznych. Transport substancji przez błony biologiczne
Wszystkie żywe organizmy, w zależności od budowy komórki, dzielą się na trzy grupy (patrz ryc. 1):
1. Prokarioty (niejądrowe)
2. Eukarionty (jądrowe)
3. Wirusy (niekomórkowe)
Ryż. 1. Organizmy żywe
Na tej lekcji zaczniemy badać strukturę komórek organizmów eukariotycznych, do których należą rośliny, grzyby i zwierzęta. Ich komórki są największe i bardziej złożone pod względem struktury w porównaniu do komórek prokariotów.
Jak wiadomo, komórki są zdolne do niezależnej aktywności. Mogą wymieniać materię i energię z otoczeniem, a także rosnąć i rozmnażać się Struktura wewnętrzna komórki są bardzo złożone i zależą przede wszystkim od funkcji, jaką komórka pełni w organizmie wielokomórkowym.
Zasady budowy wszystkich komórek są takie same. W każdej komórce eukariotycznej można wyróżnić następujące główne części (patrz ryc. 2):
1. Zewnętrzna męmbrana, który oddziela zawartość komórki od środowiska zewnętrznego.
2. Cytoplazma z organellami.

Ryż. 2. Główne części komórki eukariotycznej
Termin „membrana” został zaproponowany około sto lat temu w odniesieniu do granic komórki, ale wraz z rozwojem mikroskopii elektronowej stało się jasne, że błona komórkowa jest częścią elementów strukturalnych komórki.
W 1959 roku J.D. Robertson sformułował hipotezę dotyczącą budowy błony elementarnej, zgodnie z którą błony komórkowe zwierząt i roślin zbudowane są według tego samego typu.
W 1972 roku zaproponowali to Singer i Nicholson, co jest obecnie powszechnie akceptowane. Według tego modelu podstawą każdej membrany jest dwuwarstwa fosfolipidów.
Fosfolipidy (związki zawierające grupę fosforanową) mają cząsteczki składające się z polarnej głowy i dwóch niepolarnych ogonów (patrz ryc. 3).

Ryż. 3. Fosfolipidy
W dwuwarstwie fosfolipidowej hydrofobowe reszty kwasów tłuszczowych są skierowane do wewnątrz, a hydrofilowe główki, w tym reszta kwasu fosforowego, są skierowane na zewnątrz (patrz ryc. 4).

Ryż. 4. Dwuwarstwa fosfolipidowa
Dwuwarstwa fosfolipidowa jest przedstawiana jako struktura dynamiczna, lipidy mogą się poruszać, zmieniając swoje położenie.
Podwójna warstwa lipidów pełni funkcję barierową błony, zapobiegając rozprzestrzenianiu się zawartości komórki i przedostawaniu się substancji toksycznych do wnętrza komórki.
Obecność błony granicznej między komórką a środowiskiem była znana na długo przed pojawieniem się mikroskopu elektronowego. Fizykochemicy zaprzeczali istnieniu błony komórkowej i wierzyli, że istnieje połączenie między żywą zawartością koloidalną a środowiskiem, ale Pfeffer (niemiecki botanik i fizjolog roślin) potwierdził jego istnienie w 1890 roku.
Na początku ubiegłego wieku Overton (brytyjski fizjolog i biolog) odkrył, że szybkość przenikania wielu substancji do czerwonych krwinek jest wprost proporcjonalna do ich rozpuszczalności w lipidach. W związku z tym naukowiec zasugerował, że membrana zawiera duża liczba lipidy i substancje w nim rozpuszczone przechodzą przez niego i trafiają na drugą stronę membrany.
W 1925 roku Gorter i Grendel (amerykańscy biolodzy) wyizolowali lipidy z błony komórkowej czerwonych krwinek. Rozprowadzili powstałe lipidy na powierzchni wody o grubości jednej cząsteczki. Okazało się, że powierzchnia zajmowana przez warstwę lipidową jest dwukrotnie większa od powierzchni samej czerwonej krwinki. Dlatego naukowcy ci doszli do wniosku, że błona komórkowa składa się nie z jednej, ale z dwóch warstw lipidów.
Dawson i Danielli (angielscy biolodzy) w 1935 roku zasugerowali, że w błonach komórkowych dwucząsteczkowa warstwa lipidów jest umieszczona pomiędzy dwiema warstwami cząsteczek białka (patrz ryc. 5).

Ryż. 5. Model membranowy zaproponowany przez Dawsona i Danielli
Wraz z pojawieniem się mikroskopu elektronowego otworzyła się możliwość zapoznania się ze strukturą membrany, a następnie odkryto, że błony komórek zwierzęcych i roślinnych wyglądają jak struktura trójwarstwowa (patrz ryc. 6).

Ryż. 6. Błona komórkowa pod mikroskopem
W 1959 roku biolog J.D. Robertson, łącząc dostępne wówczas dane, wysunął hipotezę dotyczącą budowy „błony elementarnej”, w której postulował budowę wspólną dla wszystkich błon biologicznych.
Postulaty Robertsona dotyczące budowy „błony elementarnej”
1. Wszystkie membrany mają grubość około 7,5 nm.
2. W mikroskopie elektronowym wszystkie wydają się trójwarstwowe.
3. Trójwarstwowy wygląd membrany wynika z dokładnego ułożenia białek i lipidów polarnych, jakie przewidywał model Dawsona i Danielliego – centralna dwuwarstwa lipidowa jest umieszczona pomiędzy dwiema warstwami białka.
Ta hipoteza dotycząca budowy „błony elementarnej” ulegała różnym zmianom i w 1972 roku została wysunięta Model membrany z płynną mozaiką(patrz ryc. 7), co jest obecnie powszechnie akceptowane.

Ryż. 7. Model membrany z płynną mozaiką
Cząsteczki białek zanurzone są w dwuwarstwie lipidowej błony, tworząc ruchomą mozaikę. Ze względu na lokalizację w błonie oraz sposób oddziaływania z dwuwarstwą lipidową białka można podzielić na:
- powierzchowne (lub obwodowe) białka błonowe związane z hydrofilową powierzchnią dwuwarstwy lipidowej;
- integralna (membrana) białka osadzone w obszarze hydrofobowym dwuwarstwy.
Białka integralne różnią się stopniem osadzenia w obszarze hydrofobowym dwuwarstwy. Można je całkowicie zanurzyć ( całka) lub częściowo zanurzony ( półintegralny), a także może przenikać przez membranę przez ( transbłonowy).
Białka błonowe można podzielić na dwie grupy ze względu na ich funkcję:
- strukturalny białka. Wchodzą w skład błon komórkowych i uczestniczą w utrzymaniu ich struktury.
- dynamiczny białka. Znajdują się na membranach i biorą udział w procesach na nich zachodzących.
Istnieją trzy klasy białek dynamicznych.
1. Chwytnik. Za pomocą tych białek komórka odczuwa różne wpływy na swojej powierzchni. Oznacza to, że specyficznie wiążą związki takie jak hormony, neuroprzekaźniki i toksyny na zewnątrz błony, co służy jako sygnał do zmiany różnych procesów wewnątrz komórki lub samej błony.
2. Transport. Białka te transportują pewne substancje przez błonę, a także tworzą kanały, przez które różne jony są transportowane do i z komórki.
3. Enzymatyczny. Są to białka enzymatyczne, które znajdują się w błonie i biorą udział w różnych procesach chemicznych.
Transport substancji przez membranę
Dwuwarstwy lipidowe są w dużej mierze nieprzepuszczalne dla wielu substancji, dlatego do transportu substancji przez błonę potrzebna jest duża ilość energii, a także wymagane jest tworzenie różnych struktur.
Istnieją dwa rodzaje transportu: pasywny i aktywny.
Transport pasywny
Transport pasywny polega na przenoszeniu cząsteczek zgodnie z gradientem stężeń. Oznacza to, że zależy to jedynie od różnicy stężenia przenoszonej substancji po przeciwnych stronach membrany i odbywa się bez wydatku energetycznego.
Istnieją dwa rodzaje transportu pasywnego:
- prosta dyfuzja(patrz ryc. 8), co zachodzi bez udziału białka błonowego. Mechanizm dyfuzji prostej polega na transbłonowym przenoszeniu gazów (tlenu i dwutlenku węgla), wody i niektórych prostych jonów organicznych. Prosta dyfuzja ma niski współczynnik.

Ryż. 8. Prosta dyfuzja
- ułatwiona dyfuzja(patrz ryc. 9) różni się od prostego tym, że zachodzi przy udziale białek nośnikowych. Proces ten jest specyficzny i zachodzi z większą szybkością niż zwykła dyfuzja.

Ryż. 9. Ułatwiona dyfuzja
Znane są dwa typy białek transportujących błonę: białka nośnikowe (translokazy) i białka tworzące kanały. Białka transportowe wiążą określone substancje i transportują je przez błonę zgodnie z gradientem stężeń, dzięki czemu proces ten, podobnie jak przy prostej dyfuzji, nie wymaga wydatku energii ATP.
Cząsteczki jedzenia nie mogą przejść przez błonę, dostają się do komórki na drodze endocytozy (patrz ryc. 10). Podczas endocytozy błona plazmatyczna tworzy wgłębienia i wypustki oraz wychwytuje stałe cząstki pokarmu. Wokół bolusa pokarmowego tworzy się wakuola (lub pęcherzyk), która następnie oddziela się od błony komórkowej, a cząstki stałe w wakuoli trafiają do wnętrza komórki.

Ryż. 10. Endocytoza
Istnieją dwa rodzaje endocytozy.
1. Fagocytoza- absorpcja cząstek stałych. Nazywa się wyspecjalizowane komórki przeprowadzające fagocytozę fagocyty.
2. Pinocytoza- absorpcja materiału ciekłego (roztwór, roztwór koloidalny, zawiesina).
Egzocytoza(patrz ryc. 11) jest procesem odwrotnym do endocytozy. Substancje syntetyzowane w komórce, takie jak hormony, pakowane są w pęcherzyki błonowe, które pasują do błony komórkowej, są w niej osadzane, a zawartość pęcherzyka jest uwalniana z komórki. W ten sam sposób komórka może pozbyć się zbędnych produktów przemiany materii.

Ryż. 11. Egzocytoza
Transport aktywny
W przeciwieństwie do dyfuzji ułatwionej, transport aktywny polega na przemieszczaniu się substancji wbrew gradientowi stężeń. W tym przypadku substancje przemieszczają się z obszaru o niższym stężeniu do obszaru o wyższym stężeniu. Ponieważ ruch ten zachodzi w kierunku przeciwnym do normalnej dyfuzji, komórka musi zużywać w tym procesie energię.
Spośród przykładów transportu aktywnego najlepiej zbadaną jest tzw. pompa sodowo-potasowa. Pompa ta wypompowuje jony sodu z komórki i pompuje jony potasu do komórki, wykorzystując energię ATP.
1. Strukturalny (błona komórkowa oddziela komórkę od środowisko).
2. Transport (substancje transportowane są przez błonę komórkową, a błona komórkowa jest filtrem wysoce selektywnym).
3. Receptor (receptory zlokalizowane na powierzchni błony odbierają wpływy zewnętrzne i przekazują tę informację do wnętrza komórki, pozwalając jej szybko reagować na zmiany w otoczeniu).
Oprócz powyższego membrana pełni także funkcje metaboliczne i przekształcające energię.
Funkcja metaboliczna
Błony biologiczne bezpośrednio lub pośrednio uczestniczą w procesach przemian metabolicznych substancji w komórce, ponieważ większość enzymów jest związana z błonami.
Środowisko lipidowe enzymów w błonie stwarza określone warunki ich funkcjonowania, nakłada ograniczenia na aktywność białek błonowych, a tym samym wpływa regulująco na procesy metaboliczne.
Funkcja konwersji energii
Najważniejszą funkcją wielu biomembran jest konwersja jednej formy energii w inną.
Błony przetwarzające energię obejmują błony wewnętrzne mitochondriów i tylakoidy chloroplastów (patrz ryc. 12).

Ryż. 12. Mitochondria i chloroplasty
Bibliografia
- Kamensky A.A., Kriksunov E.A., Pasechnik V.V. Biologia ogólna Drop w klasie 10-11, 2005.
- Biologia. klasa 10. Biologia ogólna. Podstawowy poziom/ P.V. Iżewski, O.A. Kornilova, T.E. Loschilina i inni - wyd. 2, poprawione. - Ventana-Graf, 2010. - 224 s.
- Belyaev D.K. Biologia w klasie 10-11. Biologia ogólna. Podstawowy poziom. - wyd. 11, stereotyp. - M.: Edukacja, 2012. - 304 s.
- Agafonova I.B., Zakharova E.T., Sivoglazov V.I. Biologia w klasie 10-11. Biologia ogólna. Podstawowy poziom. - wyd. 6, dod. - Drop, 2010. - 384 s.
- Ayzdorov.ru ().
- Youtube.com().
- Doctor-v.ru ().
- Zwierzęta-świat.ru ().
Praca domowa
- Jaka jest budowa błony komórkowej?
- Dzięki jakim właściwościom lipidy mogą tworzyć błony?
- Dzięki jakim funkcjom białka mogą brać udział w transporcie substancji przez błonę?
- Wymień funkcje błony komórkowej.
- Jak zachodzi transport bierny przez błonę?
- Jak zachodzi transport aktywny przez błonę?
- Jaka jest funkcja pompy sodowo-potasowej?
- Co to jest fagocytoza, pinocytoza?
Krótki opis:
Sazonov V.F. 1_1 Struktura błony komórkowej [Zasoby elektroniczne] // Kinezjolog, 2009-2018: [strona internetowa]. Data aktualizacji: 02.06.2018..__.201_). _Opisano budowę i funkcjonowanie błony komórkowej (synonimy: plazmalema, plazmalemma, biomembrana, błona komórkowa, zewnętrzna błona komórkowa, błona komórkowa, błona cytoplazmatyczna). Ta wstępna informacja jest niezbędna zarówno do cytologii, jak i do zrozumienia procesów aktywność nerwowa: pobudzenie nerwowe, hamowanie, praca synaps i receptorów czuciowych.
Błona komórkowa (osocze) A lemat lub plazma O lemat)
Definicja pojęcia
Błona komórkowa(synonimy: plazmalema, plazmalemma, błona cytoplazmatyczna, biomembrana) to potrójna membrana lipoproteinowa (tj. „białko tłuszczowe”), która oddziela komórkę od środowiska i zapewnia kontrolowaną wymianę i komunikację między komórką a jej otoczeniem.
Najważniejsze w tej definicji nie jest to, że błona oddziela komórkę od środowiska, ale właśnie to łączy komórka z otoczeniem. Membrana jest aktywny struktura komórki, ona stale pracuje.
Błona biologiczna to ultracienka dwumolekularna warstwa fosfolipidów inkrustowanych białkami i polisacharydami. Ta struktura komórkowa leży u podstaw właściwości barierowych, mechanicznych i matrycowych żywego organizmu (Antonov V.F., 1996).
Graficzne przedstawienie membrany
Dla mnie błona komórkowa wygląda jak kratowe ogrodzenie z wieloma drzwiami, które otacza określone terytorium. Każde małe żywe stworzenie może swobodnie poruszać się tam i z powrotem przez to ogrodzenie. Jednak więksi goście mogą wejść tylko przez drzwi, a nawet wtedy nie przez wszystkie drzwi. Różni goście mają klucze tylko do swoich drzwi i nie mogą przejść przez drzwi innych osób. Tak więc przez to ogrodzenie stale przepływają goście tam i z powrotem, ponieważ główna funkcja Membrana ogrodzeniowa ma podwójne znaczenie: oddziela terytorium od otaczającej go przestrzeni i jednocześnie łączy ją z otaczającą przestrzenią. Dlatego w płocie jest wiele dziur i drzwi - !
Właściwości membrany
1. Przepuszczalność.
2. Półprzepuszczalność (częściowa przepuszczalność).
3. Selektywna (synonim: selektywna) przepuszczalność.
4. Przepuszczalność czynna (synonim: transport aktywny).
5. Kontrolowana przepuszczalność.
Jak widać, główną właściwością membrany jest jej przepuszczalność dla różnych substancji.
6. Fagocytoza i pinocytoza.
7. Egzocytoza.
8. Dostępność instalacji elektrycznej i potencjały chemiczne a dokładniej różnica potencjałów pomiędzy wewnętrzną i zewnętrzną stroną membrany. Można to powiedzieć obrazowo „membrana zamienia ogniwo w „baterię elektryczną”, kontrolując przepływ jonów”. Detale: .
9. Zmiany potencjału elektrycznego i chemicznego.
10. Drażliwość. Specjalne receptory molekularne zlokalizowane na błonie mogą łączyć się z substancjami sygnalizacyjnymi (kontrolującymi), w wyniku czego może zmienić się stan błony i całej komórki. Receptory molekularne wyzwalają bio reakcje chemiczne w odpowiedzi na połączenie z nimi ligandów (substancji kontrolnych). Warto zaznaczyć, że substancja sygnalizacyjna oddziałuje na receptor z zewnątrz, a zmiany zachodzą wewnątrz komórki. Okazuje się, że membrana przenosiła informację z otoczenia do środowiska wewnętrznego komórki.
11. Katalityczna aktywność enzymatyczna. Enzymy mogą być osadzone w błonie lub związane z jej powierzchnią (zarówno wewnątrz, jak i na zewnątrz komórki) i tam realizują swoje działania enzymatyczne.
12. Zmiana kształtu powierzchni i jej powierzchni. Dzięki temu błona może tworzyć wyrostki na zewnątrz lub odwrotnie, wgłębienia w komórce.
13. Zdolność do nawiązywania kontaktów z innymi błonami komórkowymi.
14. Przyczepność – zdolność przylegania do twardych powierzchni.
Krótka lista właściwości membrany
- Przepuszczalność.
- Endocytoza, egzocytoza, transcytoza.
- Potencjały.
- Drażliwość.
- Aktywność enzymatyczna.
- Łączność.
- Przyczepność.
Funkcje membrany
1. Niepełna izolacja treści wewnętrznych od środowiska zewnętrznego.
2. Najważniejsze w funkcjonowaniu błony komórkowej jest giełda różny Substancje pomiędzy komórką a środowiskiem międzykomórkowym. Wynika to z właściwości membrany polegającej na przepuszczalności. Dodatkowo membrana reguluje tę wymianę poprzez regulację jej przepuszczalności.
3. Inną ważną funkcją membrany jest tworząc różnicę potencjałów chemicznych i elektrycznych pomiędzy jego wewnętrzną i zewnętrzną stroną. Z tego powodu wnętrze ogniwa ma ujemny potencjał elektryczny - .
4. Membrana również działa wymiana informacji pomiędzy komórką a jej otoczeniem. Specjalne receptory molekularne zlokalizowane na błonie mogą wiązać się z substancjami kontrolnymi (hormonami, mediatorami, modulatorami) i wywoływać reakcje biochemiczne w komórce, prowadząc do różnorodnych zmian w funkcjonowaniu komórki lub jej strukturze.
Wideo:Struktura błony komórkowej
Wykład wideo:Szczegóły dotyczące struktury membrany i transportu
Struktura membrany
Błona komórkowa ma uniwersalny charakter trójwarstwowy Struktura. Jego środkowa warstwa tłuszczu jest ciągła, a górna i dolna warstwa białka pokrywają ją w formie mozaiki odrębnych obszarów białkowych. Warstwa tłuszczowa jest podstawą zapewniającą izolację komórki od środowiska, izolując ją od środowiska. Sam w sobie bardzo słabo przepuszcza substancje rozpuszczalne w wodzie, ale z łatwością przepuszcza substancje rozpuszczalne w tłuszczach. Dlatego przepuszczalność membrany dla substancji rozpuszczalnych w wodzie (na przykład jonów) musi być zapewniona przez specjalne struktury białkowe - i.
Poniżej znajdują się mikrofotografie rzeczywistych błon komórkowych stykających się komórek uzyskane za pomocą mikroskopu elektronowego, a także schematyczny rysunek przedstawiający trójwarstwową strukturę membrany i mozaikowy charakter jej warstw białkowych. Aby powiększyć obraz, kliknij na niego.

Oddzielny obraz wewnętrznej warstwy lipidowej (tłuszczowej) błony komórkowej, przesiąkniętej integralnymi osadzonymi białkami. Górną i dolną warstwę białkową usunięto, aby nie zakłócać oglądania dwuwarstwy lipidowej

Rysunek powyżej: Częściowe schematyczne przedstawienie błony komórkowej (błony komórkowej), podane w Wikipedii.
Należy pamiętać, że zewnętrzna i wewnętrzna warstwa białkowa została w tym miejscu usunięta z membrany, dzięki czemu możemy lepiej zobaczyć centralną dwuwarstwę tłuszczowo-lipidową. W prawdziwej błonie komórkowej duże „wyspy” białkowe unoszą się nad i pod filmem tłuszczowym (małe kulki na rysunku), a błona okazuje się grubsza, trójwarstwowa: białko-tłuszcz-białko . Czyli właściwie to jak kanapka złożona z dwóch białkowych „kawałków chleba” z tłustą warstwą „masła” w środku, czyli tzw. ma budowę trójwarstwową, a nie dwuwarstwową.
Na tym zdjęciu małe niebiesko-białe kulki odpowiadają hydrofilowym (zwilżalnym) „głowom” lipidów, a dołączone do nich „sznurki” odpowiadają hydrofobowym (niezwilżalnym) „ogonom”. Spośród białek pokazane są tylko integralne białka błonowe typu koniec do końca (czerwone kuleczki i żółte helisy). Żółte owalne kropki wewnątrz błony to cząsteczki cholesterolu, żółto-zielone łańcuchy koralików na zewnątrz błony to łańcuchy oligosacharydów tworzących glikokaliks. Glikokaliks to rodzaj węglowodanowego („cukrowego”) „puchu” na błonie, utworzonego przez wystające z niego długie cząsteczki węglowodanowo-białkowe.
Życie to mały „worek białkowo-tłuszczowy” wypełniony półpłynną, galaretowatą zawartością, która jest przesiąknięta foliami i rurkami.
Ściany tego worka utworzone są przez podwójną warstwę tłuszczową (lipidową), pokrytą wewnątrz i na zewnątrz białkami – błoną komórkową. Dlatego mówią, że membrana ma trójwarstwowa struktura : białka-tłuszcze-białka. Wewnątrz komórki znajduje się również wiele podobnych błon tłuszczowych, które ją dzielą przestrzeń wewnętrzna do przedziałów. Te same błony otaczają organelle komórkowe: jądro, mitochondria, chloroplasty. Zatem błona jest uniwersalną strukturą molekularną wspólną dla wszystkich komórek i wszystkich żywych organizmów.

Po lewej stronie nie widać już rzeczywistego, lecz sztuczny model fragmentu błony biologicznej: jest to chwilowa migawka dwuwarstwy tłuszczowej fosfolipidów (tj. podwójnej warstwy) w procesie jej symulacji dynamiki molekularnej. Pokazano komórkę obliczeniową modelu - 96 cząsteczek PC ( F osfatydyl X olina) i 2304 cząsteczki wody, co daje w sumie 20544 atomy.
Po prawej stronie znajduje się wizualny model pojedynczej cząsteczki tego samego lipidu, z którego zbudowana jest dwuwarstwa lipidowa błony. Na górze ma hydrofilową (lubiącą wodę) głowę, a na dole dwa hydrofobowe (bojące się wody) ogony. Lipid ten ma prostą nazwę: 1-steroilo-2-dokozaheksaenoilo-Sn-glicero-3-fosfatydylocholina (18:0/22:6(n-3)cis PC), ale nie musisz o tym pamiętać, chyba że planujesz sprawić, że twój nauczyciel zemdleje głębią swojej wiedzy.
Można podać dokładniejsze definicja naukowa klatka szybowa:
to uporządkowany, ustrukturyzowany, heterogeniczny układ biopolimerów otoczony aktywną membraną, uczestniczący w jednym zestawie procesów metabolicznych, energetycznych i informacyjnych, a także utrzymujący i odtwarzający cały system jako całość.
Wnętrze komórki również jest przesiąknięte błonami, a pomiędzy błonami nie znajduje się woda, lecz lepki żel/zol o zmiennej gęstości. Dlatego oddziałujące cząsteczki w komórce nie unoszą się swobodnie, jak w probówce z roztworem wodnym, ale przeważnie osiadają (unieruchomione) na strukturach polimerowych cytoszkieletu lub błonach wewnątrzkomórkowych. Dlatego reakcje chemiczne zachodzą wewnątrz komórki prawie tak, jak w ciele stałym, a nie w cieczy. Zewnętrzna błona otaczająca komórkę jest również wyłożona enzymami i receptorami molekularnymi, co czyni ją bardzo aktywną częścią komórki.
Błona komórkowa (plazmalemma, plazmolemma) jest aktywną membraną, która oddziela komórkę od środowiska i łączy ją z otoczeniem. © Sazonov V.F., 2016.
Z tej definicji membrany wynika, że nie tylko ogranicza ona komórkę, ale także aktywnie pracujełącząc go z otoczeniem.
Tłuszcz tworzący błony jest wyjątkowy, dlatego jego cząsteczki zwykle nazywane są nie tylko tłuszczem, ale „lipidy”, „fosfolipidy”, „sfingolipidy”. Folia membranowa jest podwójna, to znaczy składa się z dwóch sklejonych ze sobą folii. Dlatego w podręcznikach piszą, że podstawa błony komórkowej składa się z dwóch warstw lipidowych (lub „ dwuwarstwowy", czyli podwójną warstwę). Dla każdej pojedynczej warstwy lipidowej jedną stronę można zwilżyć wodą, drugiej nie. Dzięki temu folie te przylegają do siebie dokładnie swoimi niezwilżalnymi stronami.
Błona bakteryjna
Ściana komórkowa prokariotyczna bakterii Gram-ujemnych składa się z kilku warstw, jak pokazano na poniższym rysunku.
Warstwy otoczki bakterii Gram-ujemnych:
1. Wewnętrzna trójwarstwowa błona cytoplazmatyczna, która styka się z cytoplazmą.
2. Ściana komórkowa zbudowana z mureiny.
3. Zewnętrzna trójwarstwowa błona cytoplazmatyczna, która ma ten sam układ lipidów z kompleksami białkowymi, co błona wewnętrzna.
Komunikacja komórek bakterii Gram-ujemnych z świat zewnętrzny dzięki tak złożonej, trójstopniowej strukturze nie daje im przewagi w przetrwaniu w trudnych warunkach w porównaniu z bakteriami Gram-dodatnimi, które mają słabszą otoczkę. Nie tolerują też wysokich temperatur, zwiększonej kwasowości i zmian ciśnienia.




Wykład wideo:Membrana plazmowa. E.V. Cheval, dr.
Wykład wideo:Błona jako granica komórki. A. Iliaskin
Znaczenie kanałów jonowych membrany
Łatwo zrozumieć, że tylko substancje rozpuszczalne w tłuszczach mogą przedostać się do komórki przez błonę tłuszczową. Są to tłuszcze, alkohole, gazy. Na przykład w czerwonych krwinkach tlen i dwutlenek węgla łatwo przedostają się i wypływają bezpośrednio przez błonę. Ale woda i substancje rozpuszczalne w wodzie (na przykład jony) po prostu nie mogą przejść przez membranę do żadnej komórki. Oznacza to, że wymagają specjalnych otworów. Ale jeśli po prostu zrobisz dziurę w warstwie tłuszczu, natychmiast się ona zamknie. Co robić? Rozwiązanie znaleziono w naturze: konieczne jest wykonanie specjalnych struktur transportujących białka i rozciągnięcie ich przez membranę. Dokładnie w ten sposób powstają kanały dla przejścia substancji nierozpuszczalnych w tłuszczach - kanały jonowe błony komórkowej.
Zatem, aby nadać swojej błonie dodatkowe właściwości przepuszczalności cząsteczek polarnych (jonów i wody), komórka syntetyzuje w cytoplazmie specjalne białka, które następnie integrują się z błoną. Występują w dwóch rodzajach: białka transportowe (na przykład ATPazy transportowe) i białka tworzące kanały (twórcy kanałów). Białka te osadzone są w podwójnej warstwie tłuszczowej błony i tworzą struktury transportowe w postaci transporterów lub w postaci kanałów jonowych. Różne substancje rozpuszczalne w wodzie, które w przeciwnym razie nie mogłyby przejść przez błonę tłuszczową, mogą teraz przejść przez te struktury transportowe.
Ogólnie nazywane są również białka osadzone w błonie całka, właśnie dlatego, że wydają się być zawarte w membranie i przez nią przenikają. Inne białka, nie integralne, tworzą jakby wyspy „unoszące się” na powierzchni błony: albo na jej zewnętrznej, albo na wewnętrznej powierzchni. W końcu każdy wie, że tłuszcz to dobry smar i łatwo się po nim ślizgać!
wnioski
1. Ogólnie membrana okazuje się trójwarstwowa:
1) zewnętrzna warstwa z białkowych „wysp”
2) tłuszczowe dwuwarstwowe „morze” (dwuwarstwa lipidowa), tj. podwójny film lipidowy,
3) wewnętrzna warstwa „wysp” białkowych.
Ale istnieje również luźna warstwa zewnętrzna - glikokaliks, który jest utworzony przez glikoproteiny wystające z błony. Są to receptory molekularne, z którymi wiążą się substancje kontrolujące sygnalizację.
2. W membranę wbudowane są specjalne struktury białkowe, zapewniające jej przepuszczalność dla jonów lub innych substancji. Nie wolno nam zapominać, że w niektórych miejscach morze tłuszczu jest przesiąknięte integralnymi białkami. I to właśnie białka integralne tworzą coś wyjątkowego konstrukcje transportowe błona komórkowa (patrz rozdział 1_2 Mechanizmy transportu przez błonę). Przez nie substancje dostają się do komórki, a także są usuwane z komórki na zewnątrz.
3. Po dowolnej stronie błony (zewnętrznej i wewnętrznej), a także wewnątrz błony mogą znajdować się białka enzymatyczne, które wpływają zarówno na stan samej błony, jak i na życie całej komórki.
Błona komórkowa jest zatem aktywną, zmienną strukturą, która aktywnie działa na rzecz całej komórki i łączy ją ze światem zewnętrznym, a nie jest tylko „powłoką ochronną”. To najważniejsza rzecz, którą musisz wiedzieć o błonie komórkowej.
W medycynie białka błonowe są często wykorzystywane jako „cele”. leki. Takie cele obejmują receptory, kanały jonowe, enzymy i systemy transportowe. Ostatnio oprócz błony celem leków stają się także geny ukryte w jądrze komórkowym.
Wideo:Wprowadzenie do biofizyki błony komórkowej: Struktura błony 1 (Vladimirov Yu.A.)
Wideo:Historia, budowa i funkcje błony komórkowej: Struktura membrany 2 (Vladimirov Yu.A.)
© 2010-2018 Sazonov V.F., © 2010-2016 kineziolog.bodhy.
Komórka- samoregulująca się strukturalna i funkcjonalna jednostka tkanek i narządów. Teoria komórki strukturę narządów i tkanek opracowali Schleiden i Schwann w 1839 r. Następnie za pomocą mikroskopii elektronowej i ultrawirowania udało się określić strukturę wszystkich głównych organelli komórek zwierzęcych i roślinnych (ryc. 1).
Ryż. 1. Schemat budowy komórki zwierzęcej
Głównymi częściami komórki są cytoplazma i jądro. Każda komórka jest otoczona bardzo cienką błoną, która ogranicza jej zawartość.
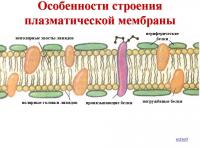
Nazywa się błona komórkowa błona plazmatyczna i charakteryzuje się selektywną przepuszczalnością. Ta właściwość pozwala na dostarczenie niezbędnych składników odżywczych i pierwiastki chemiczne wnikają do komórki, a nadmiar produktów ją opuszcza. Błona plazmatyczna składa się z dwóch warstw cząsteczek lipidów zawierających określone białka. Głównymi lipidami błonowymi są fosfolipidy. Zawierają fosfor, polarną głowę i dwa niepolarne ogony długołańcuchowych kwasów tłuszczowych. Do lipidów błonowych zalicza się cholesterol i estry cholesterolu. Zgodnie z modelem struktury płynnej mozaiki, membrany zawierają wtrącenia cząsteczek białek i lipidów, które mogą mieszać się względem dwuwarstwy. Każdy rodzaj błony dowolnej komórki zwierzęcej ma swój własny, stosunkowo stały skład lipidów.
Białka błonowe dzielą się na dwa typy w zależności od ich budowy: integralne i obwodowe. Białka obwodowe można usunąć z membrany bez jej niszczenia. Istnieją cztery rodzaje białek błonowych: białka transportowe, enzymy, receptory i białka strukturalne. Niektóre białka błonowe wykazują aktywność enzymatyczną, inne wiążą pewne substancje i ułatwiają ich transport do komórki. Białka zapewniają kilka ścieżek przemieszczania się substancji przez błony: tworzą duże pory składające się z kilku podjednostek białkowych, które umożliwiają cząsteczkom wody i jonom przemieszczanie się między komórkami; tworzą kanały jonowe wyspecjalizowane w przemieszczaniu się określonych typów jonów przez membranę w określonych warunkach. Białka strukturalne są związane z wewnętrzną warstwą lipidową i stanowią cytoszkielet komórki. Cytoszkielet zapewnia wytrzymałość mechaniczną błonie komórkowej. W różnych błonach białka stanowią od 20 do 80% masy. Białka błonowe mogą swobodnie poruszać się w płaszczyźnie bocznej.
Błona zawiera również węglowodany, które mogą być kowalencyjnie związane z lipidami lub białkami. Istnieją trzy rodzaje węglowodanów błonowych: glikolipidy (gangliozydy), glikoproteiny i proteoglikany. Większość lipidów błonowych występuje w stan ciekły i mają pewną płynność, tj. możliwość przemieszczania się z jednego obszaru do drugiego. Po zewnętrznej stronie błony znajdują się miejsca receptorowe, które wiążą różne hormony. Inne specyficzne obszary błony nie są w stanie rozpoznać i związać pewnych białek i różnych biologicznie aktywnych związków, które są obce tym komórkom.
Wewnętrzna przestrzeń komórki wypełniona jest cytoplazmą, w której zachodzi większość enzymatycznie katalizowanych reakcji metabolizmu komórkowego. Cytoplazma składa się z dwóch warstw: wewnętrznej, zwanej endoplazmą, i obwodowej, ektoplazmy, która charakteryzuje się dużą lepkością i jest pozbawiona ziarnistości. Cytoplazma zawiera wszystkie składniki komórki lub organelli. Najważniejszymi organellami komórkowymi są siateczka śródplazmatyczna, rybosomy, mitochondria, aparat Golgiego, lizosomy, mikrofilamenty i mikrotubule, peroksysomy.
Siateczka endoplazmatyczna to system połączonych ze sobą kanałów i wnęk, które przenikają całą cytoplazmę. Zapewnia transport substancji ze środowiska i wnętrza komórek. Siateczka śródplazmatyczna służy również jako magazyn wewnątrzkomórkowych jonów Ca 2+ i służy jako główne miejsce syntezy lipidów w komórce.
Rybosomy - mikroskopijne kuliste cząstki o średnicy 10-25 nm. Rybosomy są swobodnie umiejscowione w cytoplazmie lub przyczepione do zewnętrznej powierzchni błon retikulum endoplazmatycznego i błona jądrowa. Oddziałują z informacyjnym i transportowym RNA, zachodzi w nich synteza białek. Syntetyzują białka, które dostają się do cystern lub aparatu Golgiego, a następnie są uwalniane na zewnątrz. Rybosomy, swobodnie zlokalizowane w cytoplazmie, syntetyzują białko do wykorzystania przez samą komórkę, a rybosomy związane z retikulum endoplazmatycznym wytwarzają białko, które jest wydalane z komórki. Rybosomy syntetyzują różne białka funkcjonalne: białka nośnikowe, enzymy, receptory, białka cytoszkieletowe.
Aparat Golgiego utworzone przez system kanalików, cystern i pęcherzyków. Jest związany z retikulum endoplazmatycznym, a substancje biologicznie czynne, które tu dostają, są magazynowane w postaci zwartej w pęcherzykach wydzielniczych. Te ostatnie są stale oddzielane od aparatu Golgiego, transportowane do błony komórkowej i łączą się z nią, a substancje zawarte w pęcherzykach są usuwane z komórki w procesie egzocytozy.
Lizosomy - cząsteczki otoczone membraną o wielkości 0,25-0,8 mikrona. Zawierają liczne enzymy biorące udział w rozkładaniu białek, polisacharydów, tłuszczów, kwasów nukleinowych, bakterii i komórek.
Peroksysomy utworzone z gładkiej siateczki śródplazmatycznej, przypominają lizosomy i zawierają enzymy katalizujące rozkład nadtlenku wodoru, który rozkłada się pod wpływem peroksydaz i katalazy.
Mitochondria zawierają błonę zewnętrzną i wewnętrzną i są „stacją energetyczną” komórki. Mitochondria to okrągłe lub wydłużone struktury z podwójną błoną. Błona wewnętrzna tworzy fałdy wystające do mitochondriów - cristae. Zachodzi w nich synteza ATP, utlenianie substratów cyklu Krebsa i zachodzi wiele reakcji biochemicznych. Cząsteczki ATP wytwarzane w mitochondriach dyfundują do wszystkich części komórki. Mitochondria zawierają niewielką ilość DNA, RNA i rybosomów i przy ich udziale następuje odnowa i synteza nowych mitochondriów.
Mikrofilamenty Są to cienkie włókna białkowe składające się z miozyny i aktyny, tworzące aparat kurczliwy komórki. Mikrofilamenty biorą udział w tworzeniu fałd lub wypukłości błony komórkowej, a także w ruchu różnych struktur w komórkach.
Mikrotubule stanowią podstawę cytoszkieletu i zapewniają jego wytrzymałość. Cytoszkielet nadaje komórkom ich charakterystykę wygląd i kształt, służy jako miejsce przyłączania organelli wewnątrzkomórkowych i różnych ciał. W komórkach nerwowych wiązki mikrotubul biorą udział w transporcie substancji z ciała komórki do końcówek aksonów. Przy ich udziale wrzeciono mitotyczne funkcjonuje podczas podziału komórki. Pełnią rolę elementów motorycznych kosmków i wici u eukariontów.
Rdzeń jest główną strukturą komórki, uczestniczy w przekazywaniu cech dziedzicznych oraz w syntezie białek. Jądro otoczone jest błoną jądrową zawierającą wiele porów jądrowych, przez które następuje wymiana różnych substancji pomiędzy jądrem a cytoplazmą. Wewnątrz znajduje się jąderko. Ustalono ważną rolę jąderka w syntezie rybosomalnego RNA i białek histonowych. Pozostałe części jądra zawierają chromatynę składającą się z DNA, RNA i szeregu specyficznych białek.
Funkcje błony komórkowej
Błony komórkowe odgrywają kluczową rolę w regulacji metabolizmu wewnątrzkomórkowego i międzykomórkowego. Mają selektywną przepuszczalność. Ich specyficzna budowa pozwala im pełnić funkcje barierowe, transportowe i regulacyjne.
Funkcja bariery objawia się ograniczeniem przenikania przez membranę związków rozpuszczonych w wodzie. Membrana jest nieprzepuszczalna dla dużych cząsteczek białka i anionów organicznych.
Funkcja regulacyjna Celem błon komórkowych jest regulacja metabolizmu wewnątrzkomórkowego w odpowiedzi na wpływy chemiczne, biologiczne i mechaniczne. Specjalne receptory błonowe odbierają różne wpływy, co powoduje późniejszą zmianę aktywności enzymu.
Funkcja transportowa przez błony biologiczne może odbywać się w sposób pasywny (dyfuzja, filtracja, osmoza) lub przy wykorzystaniu transportu aktywnego.
Dyfuzja - ruch gazu lub substancji rozpuszczalnej wzdłuż gradientu stężenia i elektrochemicznego. Szybkość dyfuzji zależy od przepuszczalności błony komórkowej, a także gradientu stężenia dla cząstek nienaładowanych oraz gradientu elektrycznego i stężenia dla cząstek naładowanych. Prosta dyfuzja zachodzi poprzez dwuwarstwę lipidową lub kanały. Cząstki naładowane poruszają się zgodnie z gradientem elektrochemicznym, a cząstki nienaładowane poruszają się zgodnie z gradientem chemicznym. Na przykład tlen, hormony steroidowe, mocznik, alkohol itp. przenikają przez warstwę lipidową membrany na drodze prostej dyfuzji. Przez kanały przemieszczają się różne jony i cząstki. Kanały jonowe są utworzone przez białka i dzielą się na kanały bramkowane i niebramkowane. W zależności od selektywności rozróżnia się kable jonoselektywne, które przepuszczają tylko jeden jon, oraz kanały, które nie mają selektywności. Kanały posiadają kryzę i filtr selektywny, a kanały sterowane posiadają mechanizm bramkowy.
Ułatwiona dyfuzja - proces, w którym substancje są transportowane przez błonę przy użyciu specjalnych białek transportujących przez błonę. W ten sposób aminokwasy i monosacharydy przedostają się do wnętrza komórki. Ten rodzaj transportu odbywa się bardzo szybko.
Osmoza - przepływ wody przez membranę z roztworu o niższym ciśnieniu osmotycznym do roztworu o wyższym ciśnieniu osmotycznym.
Transport aktywny - transport substancji wbrew gradientowi stężeń za pomocą ATPaz transportowych (pompy jonowe). Transfer ten następuje wraz z wydatkiem energii.
W większym stopniu zbadano pompy Na + /K + -, Ca 2+ - i H + -. Pompy znajdują się na błonach komórkowych.
Rodzaj transportu aktywnego to endocytoza I egzocytoza. Dzięki tym mechanizmom transportowane są większe substancje (białka, polisacharydy, kwasy nukleinowe), które nie mogą być transportowane kanałami. Transport ten występuje częściej w komórkach nabłonka jelit, kanalikach nerkowych i śródbłonku naczyń.
Na W endocytozie błony komórkowe tworzą wgłębienia w komórce, które po uwolnieniu zamieniają się w pęcherzyki. Podczas egzocytozy pęcherzyki wraz z zawartością przedostają się do błony komórkowej i łączą się z nią, a zawartość pęcherzyków uwalniana jest do środowiska zewnątrzkomórkowego.
Budowa i funkcje błony komórkowej
Aby zrozumieć procesy zapewniające istnienie potencjałów elektrycznych w żywych komórkach, należy najpierw zrozumieć strukturę błony komórkowej i jej właściwości.
Obecnie najszerzej akceptowanym jest model ciekłej mozaiki membrany, zaproponowany przez S. Singera i G. Nicholsona w 1972 roku. Membrana zbudowana jest z podwójnej warstwy fosfolipidów (dwuwarstwy), której hydrofobowe fragmenty cząsteczki zanurzone w grubości membrany, a polarne grupy hydrofilowe są zorientowane na zewnątrz, tj. do otaczającego środowiska wodnego (ryc. 2).
Białka błonowe są zlokalizowane na powierzchni błony lub mogą być osadzone na różnej głębokości w strefie hydrofobowej. Niektóre białka rozciągają się przez błonę, a po obu stronach błony komórkowej znajdują się różne grupy hydrofilowe tego samego białka. Białka występujące w błona plazmatyczna, odgrywają bardzo ważną rolę: biorą udział w tworzeniu kanałów jonowych, pełnią rolę pomp membranowych i transporterów różne substancje, a także może pełnić funkcję receptorową.
Główne funkcje błony komórkowej: barierowa, transportowa, regulacyjna, katalityczna.
Funkcja bariery polega na ograniczeniu dyfuzji związków rozpuszczalnych w wodzie przez błonę, co jest niezbędne do ochrony komórek przed obcymi, toksycznymi substancjami i utrzymania w miarę stałej zawartości różnych substancji wewnątrz komórek. W ten sposób błona komórkowa może spowolnić dyfuzję różnych substancji od 100 000 do 10 000 000 razy.

Ryż. 2. Trójwymiarowy diagram modelu płynnej mozaiki membrany Singera-Nicholsona
Pokazano kuliste białka integralne osadzone w dwuwarstwie lipidowej. Niektóre białka są kanałami jonowymi, inne (glikoproteiny) zawierają boczne łańcuchy oligosacharydowe, które biorą udział w rozpoznawaniu komórek między sobą oraz w tkance międzykomórkowej. Cząsteczki cholesterolu przylegają ściśle do głów fosfolipidów i mocują sąsiednie odcinki „ogonów”. Wewnętrzne odcinki ogonów cząsteczki fosfolipidów nie są ograniczone w swoim ruchu i odpowiadają za płynność błony (Bretscher, 1985)
W membranie znajdują się kanały, przez które przenikają jony. Kanały mogą być zależne od napięcia lub niezależne od potencjału. Kanały zależne od napięcia otwierają się, gdy zmienia się różnica potencjałów, oraz potencjalnie niezależny(regulowane przez hormony) otwierają się, gdy receptory wchodzą w interakcję z substancjami. Kanały można otwierać i zamykać dzięki bramkom. W membranę wbudowane są dwa rodzaje bramek: aktywacja(głęboko w kanale) i inaktywacja(na powierzchni kanału). Brama może znajdować się w jednym z trzech stanów:
- stan otwarty (oba typy bramek są otwarte);
- stan zamknięty (brama aktywacji zamknięta);
- stan inaktywacji (bramka inaktywacji zamknięta).
Inny cecha charakterystyczna błony to zdolność do selektywnego transportu jonów nieorganicznych, składników odżywczych i różne produkty giełda. Istnieją systemy pasywnego i aktywnego przenoszenia (transportu) substancji. Bierny transport odbywa się poprzez kanały jonowe z udziałem białek nośnikowych lub bez nich siła napędowa jest różnicą potencjałów elektrochemicznych jonów pomiędzy przestrzenią wewnątrz- i zewnątrzkomórkową. Selektywność kanałów jonowych zależy od ich parametrów geometrycznych i Natura chemiczna grupy wyścielające ściany kanału i jego ujścia.
Obecnie najlepiej zbadanymi kanałami są te, które są selektywnie przepuszczalne dla jonów Na+, K+, Ca 2+, a także dla wody (tzw. akwaporyny). Według różnych badań średnica kanałów jonowych wynosi 0,5-0,7 nm. Pojemność kanału może być różna; przez jeden kanał jonowy może przejść 10 7 - 10 8 jonów na sekundę.
Aktywny transport odbywa się z wydatkiem energii i odbywa się za pomocą tzw. pomp jonowych. Pompy jonowe to molekularne struktury białkowe osadzone w membranie, które transportują jony w kierunku wyższego potencjału elektrochemicznego.
Pompy działają wykorzystując energię hydrolizy ATP. Obecnie Na+/K+ - ATPaza, Ca 2+ - ATPaza, H + - ATPaza, H + /K + - ATPaza, Mg 2+ - ATPaza, które zapewniają ruch odpowiednio jonów Na +, K +, Ca 2+ , zostały dobrze zbadane, H+, Mg 2+ izolowane lub sprzężone (Na+ i K+; H+ i K+). Molekularny mechanizm aktywnego transportu nie jest w pełni poznany.
Błony to niezwykle lepkie i jednocześnie plastyczne struktury otaczające wszystkie żywe komórki. Funkcje błony komórkowe:
1. Błona plazmatyczna jest barierą utrzymującą odmienny skład środowiska zewnątrz- i wewnątrzkomórkowego.
2.Błony tworzą wyspecjalizowane przedziały wewnątrz komórki, tj. liczne organelle - mitochondria, lizosomy, kompleks Golgiego, retikulum endoplazmatyczne, błony jądrowe.
3. W błonach zlokalizowane są enzymy biorące udział w konwersji energii w procesach takich jak fosforylacja oksydacyjna i fotosynteza.
Struktura i skład membran
Podstawą błony jest podwójna warstwa lipidowa, w której tworzeniu biorą udział fosfolipidy i glikolipidy. Dwuwarstwę lipidową tworzą dwa rzędy lipidów, których rodniki hydrofobowe są ukryte do wewnątrz, a grupy hydrofilowe skierowane są na zewnątrz i stykają się ze środowiskiem wodnym. Cząsteczki białek są niejako „rozpuszczone” w dwuwarstwie lipidowej.
Struktura lipidów błonowych
Lipidy błonowe są cząsteczkami amfifilowymi, ponieważ cząsteczka ma zarówno obszar hydrofilowy (głowy polarne), jak i obszar hydrofobowy, reprezentowany przez rodniki węglowodorowe kwasów tłuszczowych, które samorzutnie tworzą dwuwarstwę. Błony zawierają trzy główne typy lipidów – fosfolipidy, glikolipidy i cholesterol.
Skład lipidów jest inny. Zawartość konkretnego lipidu jest najwyraźniej zdeterminowana różnorodnością funkcji pełnionych przez te lipidy w błonach.
Fosfolipidy. Wszystkie fosfolipidy można podzielić na dwie grupy – glicerofosfolipidy i sfingofosfolipidy. Glicerofosfolipidy zaliczane są do pochodnych kwasu fosfatydowego. Najbardziej powszechnymi glicerofosfolipidami są fosfatydylocholiny i fosfatydyloetanoloaminy. Sfingofosfolipidy oparte są na sfingozynie aminoalkoholowej.
Glikolipidy. W glikolipidach część hydrofobowa jest reprezentowana przez ceramid alkoholowy, a część hydrofilowa jest reprezentowana przez resztę węglowodanową. W zależności od długości i budowy części węglowodanowej wyróżnia się cerebrozydy i gangliozydy. Polarne „głowy” glikolipidów znajdują się na zewnętrznej powierzchni błon plazmatycznych.
Cholesterol (CS). CS jest obecny we wszystkich błonach komórek zwierzęcych. Jego cząsteczka składa się ze sztywnego hydrofobowego rdzenia i elastycznego łańcucha węglowodorowego. Pojedyncza grupa hydroksylowa w pozycji 3 to „głowa polarna”. W komórce zwierzęcej średni stosunek molowy cholesterol/fosfolipidy wynosi 0,3-0,4, ale w błonie komórkowej ten stosunek jest znacznie wyższy (0,8-0,9). Obecność cholesterolu w błonach zmniejsza ruchliwość kwasów tłuszczowych, ogranicza boczną dyfuzję lipidów i dlatego może wpływać na funkcje białek błonowych.
Właściwości membrany:
1. Selektywna przepuszczalność. Zamknięta dwuwarstwa zapewnia jedną z głównych właściwości membrany: jest nieprzepuszczalna dla większości cząsteczek rozpuszczalnych w wodzie, ponieważ nie rozpuszczają się one w jej hydrofobowym rdzeniu. Gazy takie jak tlen, CO 2 i azot mają zdolność łatwego przenikania do komórek ze względu na mały rozmiar ich cząsteczek i słabe oddziaływanie z rozpuszczalnikami. Cząsteczki o charakterze lipidowym, takie jak hormony steroidowe, również łatwo przenikają przez dwuwarstwę.
2. Płynność. Błony charakteryzują się płynnością (płynnością), zdolnością lipidów i białek do poruszania się. Możliwe są dwa rodzaje ruchów fosfolipidów: salto (w literatura naukowa zwane „flip-flop”) i dyfuzją boczną. W pierwszym przypadku przeciwstawne sobie cząsteczki fosfolipidów w warstwie dwucząsteczkowej przewracają się (lub wykonują salta) ku sobie i zamieniają miejscami w błonie, tj. to, co zewnętrzne staje się wnętrzem i odwrotnie. Takie skoki wiążą się ze zużyciem energii. Częściej obserwuje się rotacje wokół osi (rotację) i dyfuzję boczną - ruch w warstwie równoległej do powierzchni membrany. Szybkość ruchu cząsteczek zależy od mikrolepkości błon, która z kolei zależy od względnej zawartości nasyconych i nienasyconych kwasów tłuszczowych w składzie lipidów. Mikrolepkość jest mniejsza, jeśli w składzie lipidów dominują nienasycone kwasy tłuszczowe, a większa, jeśli zawartość nasyconych kwasów tłuszczowych jest wysoka.
3. Asymetria membrany. Powierzchnie tej samej błony różnią się składem lipidów, białek i węglowodanów (asymetria poprzeczna). Na przykład w warstwie zewnętrznej dominują fosfatydylocholiny, a w warstwie wewnętrznej dominują fosfatydyloetanoloaminy i fosfatydyloseryny. Składniki węglowodanowe glikoprotein i glikolipidów wydostają się na powierzchnię zewnętrzną, tworząc ciągłą strukturę zwaną glikokaliksem. Na wewnętrznej powierzchni nie ma węglowodanów. Białka - receptory hormonów znajdują się na zewnętrznej powierzchni błony komórkowej, a enzymy, które regulują - cyklaza adenylanowa, fosfolipaza C - na wewnętrznej powierzchni itp.
Białka błonowe
Fosfolipidy błonowe pełnią funkcję rozpuszczalnika dla białek błonowych, tworząc mikrośrodowisko, w którym te ostatnie mogą funkcjonować. Białka stanowią od 30 do 70% masy błon. Liczba różnych białek w błonie waha się od 6-8 w siateczce sarkoplazmatycznej do ponad 100 w błonie komórkowej. Są to enzymy, białka transportowe, białka strukturalne, antygeny, w tym antygeny głównego układu zgodności tkankowej, receptory różnych cząsteczek.
Ze względu na lokalizację w błonie białka dzielimy na integralne (częściowo lub całkowicie zanurzone w błonie) i obwodowe (znajdujące się na jej powierzchni). Niektóre białka integralne przechodzą przez błonę jednokrotnie (glikoforyna), inne wielokrotnie. Na przykład fotoreceptor siatkówki i receptor β2-adrenergiczny przekraczają dwuwarstwę 7 razy.
Białka obwodowe i domeny białek integralnych, zlokalizowane na zewnętrznej powierzchni wszystkich błon, są prawie zawsze glikozylowane. Reszty oligosacharydowe chronią białko przed proteolizą i biorą także udział w rozpoznawaniu ligandów lub adhezji.
Główne błony komórkowe:
Membrana plazmowa
Błona plazmatyczna otaczająca każdą komórkę określa jej wielkość, zapewnia transport małych i dużych cząsteczek z i do komórki oraz utrzymuje różnicę w stężeniach jonów po obu stronach membrany. Błona bierze udział w kontaktach międzykomórkowych, odbiera, wzmacnia i przekazuje sygnały ze środowiska zewnętrznego do komórki. Wiele enzymów katalizujących reakcje biochemiczne jest związanych z błoną.
Membrana nuklearna
Otoczka jądrowa składa się z zewnętrznej i wewnętrznej błony jądrowej. Otoczka jądrowa ma pory, przez które RNA przenika z jądra do cytoplazmy, oraz białka regulatorowe z cytoplazmy do jądra.
Wewnętrzna błona jądrowa zawiera specyficzne białka, które mają miejsca wiązania dla głównych polipeptydów macierzy jądrowej – laminatu A, laminatu B i laminatu C. Ważną funkcją tych białek jest rozpad błony jądrowej podczas mitozy.
Błona retikulum endoplazmatycznego (ER).
Membrana ER ma liczne fałdy i załamania. Tworzy ciągłą powierzchnię wyznaczającą wewnętrzną przestrzeń zwaną jamą ER. Szorstki ER jest związany z rybosomami, na których zachodzi synteza białek błony komórkowej, ER, aparatu Golgiego, lizosomów i wydzielanych białek. Regiony ER niezawierające rybosomów nazywane są gładkimi ER. Tutaj zachodzi końcowy etap biosyntezy cholesterolu, fosfolipidów, reakcja utleniania własnych metabolitów i substancji obcych przy udziale enzymów błonowych - cytochromu P 450, reduktazy cytochromu P 450, reduktazy cytochromu b 5 i cytochromu b 5
Aparat Golgiego
Aparat Golgiego jest ważną organellą błonową odpowiedzialną za modyfikację, gromadzenie, sortowanie i kierowanie różnych substancji do odpowiednich przedziałów wewnątrzkomórkowych, a także na zewnątrz komórki. Specyficzne enzymy kompleksu błony Golgiego, glikozylotransferazy, białka glikozylanowe przy serynie, resztach treoniny lub grupie amidowej asparaginy, dopełniające tworzenie złożonych białek - glikoprotein.
Błony mitochondrialne
Mitochondria to organelle otoczone podwójną błoną, które specjalizują się w syntezie ATP poprzez fosforylację oksydacyjną. Charakterystyczną cechą zewnętrznej błony mitochondrialnej jest zawartość dużej ilości białka porynowego, które tworzy pory w błonie. Dzięki porinie błona zewnętrzna jest swobodnie przepuszczalna dla jonów nieorganicznych, metabolitów, a nawet małych cząsteczek białek (poniżej 10 kDa). Błona zewnętrzna jest nieprzepuszczalna dla dużych białek, co pozwala mitochondriom zapobiegać przedostawaniu się białek z przestrzeni międzybłonowej do cytozolu.
Wewnętrzna błona mitochondriów charakteryzuje się dużą zawartością białek, około 70%, które pełnią głównie funkcje katalityczne i transportowe. Translokazy błonowe zapewniają selektywne przenoszenie substancji z przestrzeni międzybłonowej do matrycy i w przeciwnym kierunku, enzymy biorą udział w transporcie elektronów (łańcuch transportu elektronów) i syntezie ATP.
Błona lizosomalna
Błona lizosomowa pełni rolę „tarczy” pomiędzy aktywnymi enzymami (ponad 50), które zapewniają reakcje rozkładu białek, węglowodanów, tłuszczów, kwasów nukleinowych i innej zawartości komórkowej. Błona zawiera unikalne białka, na przykład zależną od ATP pompę protonową (pompę), która utrzymuje kwaśne środowisko (pH 5), niezbędne do działania enzymów hydrolitycznych (proteazy, lipazy), a także białka transportowe, które umożliwiają Produkty rozkładu makrocząsteczek opuszczają lizosom. Takie błony chronią je przed działaniem proteaz.
Ogólne funkcje błon biologicznych następujące:
Odgraniczają zawartość komórki od środowiska zewnętrznego i zawartość organelli od cytoplazmy.
Zapewniają transport substancji do i z komórki, z cytoplazmy do organelli i odwrotnie.
Pełnią rolę receptorów (odbierając i przetwarzając substancje chemiczne ze środowiska, rozpoznając substancje komórkowe itp.).
Są katalizatorami (zapewniają procesy chemiczne w pobliżu membrany).
Weź udział w konwersji energii.
Ogólne właściwości błon biologicznych
Bez wyjątku wszystkie błony komórkowe zbudowane są wg ogólna zasada: Są to cienkie błony lipoproteinowe składające się z podwójnej warstwy cząsteczek lipidów, w której zawarte są cząsteczki białka. W masie, w zależności od rodzaju błony, udział lipidów wynosi 25-60%, a białek 40-75%. Wiele błon zawiera węglowodany, których ilość może osiągnąć 2-10%.
Błony biologiczne bardzo selektywnie przepuszczają substancje z otaczającego roztworu. Dość łatwo przepuszczają wodę i zatrzymują większość substancji rozpuszczalnych w wodzie, a przede wszystkim substancje zjonizowane lub przenoszące ładunek elektryczny. Z tego powodu biomembrany są dobrymi izolatorami elektrycznymi w roztworach soli.
Podstawa membrany wynosi podwójna warstwa lipidowa, w tworzeniu których biorą udział fosfolipidy i glikolipidy. Dwuwarstwę lipidową tworzą dwa rzędy lipidów, których rodniki hydrofobowe są ukryte do wewnątrz, a grupy hydrofilowe skierowane są na zewnątrz i stykają się ze środowiskiem wodnym. Cząsteczki białek wydają się być „rozpuszczone” w dwuwarstwie lipidowej
Przekrój błony komórkowej
Skład lipidowy błon:
Fosfolipidy. Wszystkie fosfolipidy można podzielić na 2 grupy - glicerofosfolipidy i sfingofosfolipidy. Glicerofosfolipidy zaliczane są do pochodnych kwasu fosfatydowego. Najbardziej powszechnymi glicerofosfolipidami błonowymi są fosfatydylocholiny i fosfatydyloetanoloaminy. W błonach komórek eukariotycznych odkryto ogromną liczbę różnych fosfolipidów, które są nierównomiernie rozmieszczone w różnych błonach komórkowych. Ta nierówność dotyczy rozmieszczenia zarówno głów polarnych, jak i reszt acylowych.
Specyficznymi fosfolipidami wewnętrznej błony mitochondriów są kardiolipiny (difosfatydyloglicerole), zbudowane na bazie glicerolu i dwóch reszt kwasu fosfatydowego. Są syntetyzowane przez enzymy w wewnętrznej błonie mitochondrialnej i stanowią około 22% wszystkich fosfolipidów błonowych.
Błony plazmatyczne komórek zawierają znaczne ilości sfingomielin. Sfingomieliny zbudowane są na bazie ceramidu – acylowanego aminoalkoholu sfingozyny. Grupa polarna składa się z reszty kwasu fosforowego i choliny, etanoloaminy lub seryny. Sfingomieliny są głównymi lipidami osłonki mielinowej włókien nerwowych.
Glikolipidy. W glikolipidach część hydrofobową reprezentuje ceramid. Grupa hydrofilowa to reszta węglowodanowa połączona wiązaniem glikozydowym z grupą hydroksylową przy pierwszym atomie węgla ceramidu. Wyróżnia się je w zależności od długości i struktury części węglowodanowej cerebrozydy, zawierające resztę mono- lub oligosacharydową, oraz gangliozydy, do grupy OH przyłączony jest złożony, rozgałęziony oligosacharyd zawierający kwas N-acetyloneuraminowy (NANA).
Polarne „głowy” glikosfingolipidów znajdują się na zewnętrznej powierzchni błon plazmatycznych. Glikolipidy znajdują się w znacznych ilościach w błonach komórek mózgowych, czerwonych krwinek i komórek nabłonkowych. Gangliozydy erytrocytów różnych osobników różnią się budową łańcuchów oligosacharydowych, które wykazują właściwości antygenowe.
Cholesterol. Cholesterol występuje we wszystkich błonach komórkowych zwierząt. Jego cząsteczka składa się ze sztywnego hydrofobowego rdzenia i elastycznego łańcucha węglowodorowego, przy czym jedyną grupą hydroksylową jest „głowa polarna”.
W komórce zwierzęcej średni stosunek molowy cholesterol/fosfolipid wynosi 0,3-0,4, ale w błonie komórkowej ten stosunek jest znacznie wyższy (0,8-0,9). Obecność cholesterolu w błonach zmniejsza ruchliwość kwasów tłuszczowych, ogranicza boczną dyfuzję lipidów i białek, a zatem może wpływać na funkcje białek błonowych.
W błonach roślinnych nie ma cholesterolu, za to obecne są steroidy roślinne – sitosterol i stigmasterol.
Białka błonowe: Zwykle dzieli się je na integralne (przezbłonowe) i obwodowe. Całka białka mają rozległe obszary hydrofobowe na powierzchni i są nierozpuszczalne woda. Są one połączone z lipidami błonowymi poprzez oddziaływania hydrofobowe i częściowo zanurzone w grubości dwuwarstwy lipidowej i często przenikają do dwuwarstwy, pozostawiając powierzchnia ma stosunkowo małe obszary hydrofilowe. Oddziel te białka od membrany można osiągnąć jedynie przy użyciu detergentów, takich jak siarczan dodecylu lub sole kwasy żółciowe, które niszczą warstwę lipidową i przekształcają białko w rozpuszczalne uformować (rozpuścić), tworząc z nim skojarzenia. Wszystkie dalsze operacje Oczyszczanie białek integralnych prowadzi się także w obecności detergentów. Białka obwodowe są związane z powierzchnią dwuwarstwy lipidowej sił elektrostatycznych i można je wypłukać z membrany roztworami soli.
23. Mechanizmy przenoszenia substancji przez błony: dyfuzja prosta, pasywny symport i antyport, pierwotny transport aktywny, wtórny transport aktywny, kanały regulowane (przykłady). Transport makrocząsteczek i cząstek przez błonę. Udział błon w oddziaływaniach międzykomórkowych.
Istnieje kilka mechanizmy transportu substancji przez błonę .
Dyfuzja- przenikanie substancji przez membranę zgodnie z gradientem stężeń (z obszaru, w którym ich stężenie jest wyższe, do obszaru, w którym ich stężenie jest mniejsze). Transport rozproszony substancji (wody, jonów) odbywa się przy udziale białek błonowych, które posiadają pory molekularne, lub przy udziale fazy lipidowej (dla substancji rozpuszczalnych w tłuszczach).
Z ułatwioną dyfuzją specjalne białka transportujące przez błonę selektywnie wiążą się z jednym lub drugim jonem lub cząsteczką i transportują je przez błonę zgodnie z gradientem stężeń.
Ułatwiona dyfuzja substancji
Białka translokazy występują w błonach komórkowych. Oddziałując z konkretnym ligandem, zapewniają jego dyfuzję (transport z obszaru o większym stężeniu do obszaru o niższym stężeniu) przez membranę. W przeciwieństwie do kanałów białkowych, translokazy ulegają zmianom konformacyjnym podczas interakcji z ligandem i jego transferu przez błonę. Kinetycznie transfer substancji poprzez dyfuzję ułatwioną przypomina reakcję enzymatyczną. W przypadku translokaz występuje nasycające stężenie ligandu, przy którym wszystkie miejsca wiązania białko-ligand są zajęte, a białka działają z maksymalną prędkością Vmax. Dlatego szybkość transportu substancji na drodze dyfuzji ułatwionej zależy nie tylko od gradientu stężenia transportowanego ligandu, ale także od liczby białek nośnikowych w błonie.
Istnieją translokazy, które przenoszą tylko jedną substancję rozpuszczalną w wodzie z jednej strony membrany na drugą. Ten prosty transport nazywa się „pasywny uniport”. Przykładem uniportu jest działanie GLUT-1, translokazy transportującej glukozę przez błonę erytrocytów:
Ułatwiona dyfuzja (uniport) glukozy do erytrocytów za pomocą GLUT-1 (S - cząsteczka glukozy). Cząsteczka glukozy jest związana przez transporter na zewnętrznej powierzchni błony komórkowej. Następuje zmiana konformacyjna i środek transportera zajmowany przez glukozę otwiera się do komórki. Na skutek zmian konformacyjnych transporter traci powinowactwo do glukozy, a cząsteczka zostaje uwolniona do cytozolu komórki. Oddzielenie glukozy od transportera powoduje zmiany konformacyjne w białku i powraca ono do swojej pierwotnej „informacji”.
Niektóre translokazy mogą transportować dwie różne substancje wzdłuż gradientu stężeń w tym samym kierunku - pasywny simport lub w przeciwne kierunki -pasywny antyport .
Przykładem translokazy działającej poprzez pasywny mechanizm antyportowy jest anionowy transporter błony erytrocytów. Wewnętrzna błona mitochondrialna zawiera wiele translokaz, które pełnią pasywną antyport. W procesie takiego przeniesienia następuje równoważna wymiana jonów, ale nie zawsze równoważna wymiana ładunku.
Podstawowy transport aktywny
Transfer niektórych jonów nieorganicznych następuje wbrew gradientowi stężeń przy udziale transportowych ATPaz (pompy jonowe). Wszystkie pompy jonowe służą jednocześnie jako enzymy zdolne do autofosforylacji i autodefosforylacji. ATPazy różnią się specyficznością jonową, liczbą transportowanych jonów i kierunkiem transportu. W wyniku działania ATPazy przeniesione jony gromadzą się po jednej stronie membrany. W błonie komórkowej ludzkich komórek najczęściej występują Ma+,K+-ATPaza, Ca2+-ATPaza i H+,K+,-ATPaza błony śluzowej żołądka.
Na+, K+-ATPaza
Ten enzym transportowy katalizuje zależny od ATP transport jonów Na+ i K+ przez błonę komórkową. Ka+,K+-ATPaza składa się z podjednostek α i β; α jest dużą podjednostką katalityczną, a β jest małą podjednostką (glikoproteiną). Aktywną formą translokazy jest tetramer (αβ)2.
Na+,K+-ATPaza odpowiada za utrzymanie wysokiego stężenia K+ w komórce i niskiego stężenia Na+. Ponieważ Na+D+-ATPaza wypompowuje trzy dodatnio naładowane jony i pompuje dwa, na membranie pojawia się potencjał elektryczny o wartości ujemnej wewnątrz komórki w stosunku do jej zewnętrznej powierzchni.
Ca2+-ATPaza zlokalizowane nie tylko w błonie komórkowej, ale także w błonie ER. Enzym składa się z dziesięciu domen transbłonowych rozciągających się przez błonę komórkową. Pomiędzy drugą i trzecią domeną znajduje się kilka reszt kwasu asparaginowego zaangażowanych w wiązanie wapnia. Region pomiędzy domeną czwartą i piątą zawiera miejsce wiązania ATP i autofosforylacji reszty kwasu asparaginowego. Ca2+-ATPazy błon plazmatycznych niektórych komórek są regulowane przez białko kalmodulinę. Każda z Ca2+-ATPaz błony komórkowej i ER jest reprezentowana przez kilka izoform.
Wtórny transport aktywny
Transport niektórych substancji rozpuszczalnych wbrew gradientowi stężeń polega na jednoczesnym lub sekwencyjnym transporcie innej substancji wzdłuż gradientu stężeń w tym samym kierunku (aktywny symport) lub w kierunku przeciwnym (aktywny antyport). W komórkach ludzkich jonem transportowanym zgodnie z gradientem stężeń jest najczęściej Na+.

Kolejność zdarzeń podczas działania Ca2*-ATPazy.
1 - wiązanie dwóch jonów wapnia przez miejsce ATPazy skierowane w stronę cytozolu;
2 - zmiana ładunku i konformacji enzymu (ATPazy), spowodowana dodatkiem dwóch jonów Ca2+, prowadzi do wzrostu powinowactwa do ATP i aktywacji autofosforylacji;
3 - autofosforylacji towarzyszą zmiany informacji, ATPaza zamyka się po wewnętrznej stronie błony i otwiera się na zewnątrz;
4 - maleje powinowactwo centrów wiązania jonów wapnia i następuje ich oddzielenie od ATPazy;
5 - autodefosforylacja jest aktywowana przez jony magnezu, w wyniku czego Ca2+-ATPaza traci resztę fosforową i dwa jony Mg2+;
6 - ATPaza powraca do swojego pierwotnego stanu.
Przykładem tego rodzaju transportu jest wymiennik Na+,Ca2+ błony komórkowej (aktywny antyport), jony sodu przenoszone są wzdłuż gradientu stężeń do komórki, a jony Ca2+ wbrew gradientowi stężeń opuszczają komórkę.
Zgodnie z aktywnym mechanizmem symportu glukoza jest wchłaniana przez komórki jelitowe, a glukoza i aminokwasy są wchłaniane ponownie z moczu pierwotnego przez komórki nerek.
Transport makrocząsteczek i cząstek przez błonę: endocytoza i egzocytoza
Makrocząsteczki białek, kwasów nukleinowych, polisacharydów, kompleksów lipoproteinowych itp. nie przechodzą przez błony komórkowe, w przeciwieństwie do jonów i monomerów. Transport makrocząsteczek, ich kompleksów i cząstek do komórki odbywa się zupełnie inaczej – poprzez endocytozę. Na endocytoza (endo...- do wewnątrz) pewien obszar plazmalemy wychwytuje i niejako otacza materiał zewnątrzkomórkowy, zamykając go w wakuoli błonowej powstałej w wyniku inwazji błony. Następnie taka wakuola łączy się z lizosomem, którego enzymy rozkładają makrocząsteczki na monomery.
Odwrotny proces endocytozy jest egzocytoza (egzo...- na zewnątrz). Dzięki niemu komórka usuwa produkty wewnątrzkomórkowe lub niestrawione pozostałości zamknięte w wakuolach lub pęcherzykach. Pęcherzyk zbliża się do błony cytoplazmatycznej, łączy się z nią, a jego zawartość jest uwalniana do środowiska. W ten sposób usuwane są enzymy trawienne, hormony, hemiceluloza itp.
Zatem błony biologiczne, jako główne elementy strukturalne komórki, służą nie tylko jako granice fizyczne, ale są dynamicznymi powierzchniami funkcjonalnymi. Na błonach organelli zachodzą liczne procesy biochemiczne, takie jak aktywna absorpcja substancji, konwersja energii, synteza ATP itp.
UDZIAŁ BŁON W INTERAKCJACH MIĘDZYKOMÓRKOWYCH
Błona plazmatyczna komórek eukariotycznych zawiera wiele wyspecjalizowanych receptorów, które wchodząc w interakcję z ligandami, powodują specyficzne odpowiedzi komórkowe. Niektóre receptory wiążą cząsteczki sygnalizacyjne – hormony, neuroprzekaźniki, inne – składniki odżywcze i metabolity, jeszcze inne – uczestniczą w adhezji komórek. Do tej klasy należą receptory niezbędne do wzajemnego rozpoznawania się komórek i ich adhezji, a także receptory odpowiedzialne za wiązanie się komórek z białkami macierzy zewnątrzkomórkowej, takimi jak fibronektyna czy kolagen.
Zdolność komórek do specyficznego wzajemnego rozpoznawania się i adhezji jest ważna dla rozwoju embrionalnego. U dorosłego człowieka interakcje adhezyjne komórka-komórka i komórka-macierz są w dalszym ciągu istotne dla utrzymania stabilności tkanki. Spośród dużej rodziny receptorów adhezji komórkowej najczęściej badane są integryny, selektyny i kadheryny.
Integryny- rozległa nadrodzina homologicznych receptorów powierzchniowych komórek dla cząsteczek macierzy międzykomórkowej, takich jak kolagen, fibronektyna, laminina itp. Będąc białkami transbłonowymi, oddziałują zarówno z cząsteczkami zewnątrzkomórkowymi, jak i wewnątrzkomórkowymi białkami cytoszkieletu. Dzięki temu integryny uczestniczą w przenoszeniu informacji ze środowiska pozakomórkowego do komórki, decydując tym samym o kierunku jej różnicowania, kształcie, aktywności mitotycznej i zdolności do migracji. Informacje mogą być przekazywane także w odwrotnym kierunku – od białek wewnątrzkomórkowych poprzez receptor do macierzy zewnątrzkomórkowej.
Przykłady niektórych integryn:
receptory białek macierzy zewnątrzkomórkowej. Wiążą się ze składnikami glikoproteinowymi macierzy zewnątrzkomórkowej, w szczególności z fibronektyną, lamininą i witronektyną (patrz punkt 15);
Integryny płytkowe (IIb i IIIa) biorą udział w agregacji płytek krwi, która zachodzi podczas krzepnięcia krwi;
białka adhezyjne leukocytów. Aby migrować do miejsca zakażenia i stanu zapalnego, leukocyty muszą oddziaływać z komórkami śródbłonka naczyń. Ta interakcja może pośredniczyć w wiązaniu limfocytów T z fibroblastami podczas stanu zapalnego.
Kadheryny i selektyny - rodzina transbłonowych glikoprotein zależnych od Ca 2+ zaangażowanych w adhezję międzykomórkową. Trzy możliwe sposoby udział receptorów tego typu w adhezji międzykomórkowej.


Receptor fibronektyny. Receptor fibronektyny należy do rodziny integryn. Każda podjednostka ma pojedynczą domenę transbłonową, krótką domenę cytoplazmatyczną i rozszerzoną domenę N-zewnątrzkomórkową. Obie podjednostki (α, β) integryny są glikozylowane i utrzymywane razem za pomocą wiązań niekowalencyjnych. Podjednostka α jest syntetyzowana jako pojedynczy łańcuch polipeptydowy, następnie rozszczepiana na mały łańcuch transbłonowy i duży łańcuch zewnątrzkomórkowy, połączone mostkami dwusiarczkowymi . Podjednostka β zawiera 4 powtórzenia po 40 reszt aminokwasowych każde. Podjednostki α są bogate w cysteinę i zawierają wiele wewnątrzłańcuchowych wiązań dwusiarczkowych (niepokazane na rysunku). Wiążąc się z fibronektyną na zewnątrz i z cytoszkieletem wewnątrz komórki, integryna działa jako łącznik transbłonowy.
Metody oddziaływania cząsteczek powierzchniowych komórki w procesie adhezji międzykomórkowej. A - receptory jednej komórki mogą wiązać się z tymi samymi receptorami komórek sąsiednich (wiązanie homofilne); B - receptory jednej komórki mogą wiązać się z receptorami innego typu sąsiadujących komórek (wiązanie heterofilne); B - receptory powierzchniowe sąsiadujących komórek mogą komunikować się ze sobą za pomocą wielowartościowych cząsteczek łącznikowych.
Kadheryny z różnych tkanek są bardzo podobne, homologiczne sekwencje aminokwasów stanowią 50-60%. Każdy receptor ma jedną domenę transbłonową.
Najpełniej scharakteryzowane są trzy grupy receptorów kadheryny:
E-kadheryna występuje na powierzchni wielu komórek tkanek nabłonkowych i embrionalnych;
N-kadheryna zlokalizowana jest na powierzchni komórek nerwowych, komórek serca i komórek soczewki;
P-kadheryna znajduje się na komórkach łożyska i naskórka.
Kadheryny odgrywają ważną rolę w początkowej adhezji międzykomórkowej, na etapach morfogenezy i organogenezy, zapewniają integralność strukturalną i polarność tkanek, zwłaszcza monowarstwy nabłonkowej.
W rodzinie selektyna Trzy najlepiej zbadane białka receptorowe to L-selektyna, P-selektyna i E-selektyna. Zewnątrzkomórkowa część selektyn składa się z 3 domeny: pierwsza domena jest reprezentowana przez 2-9 bloków powtarzających się reszt aminokwasowych (białko regulatorowe dopełniacza), druga to domena naskórkowego czynnika wzrostu (EGF), trzecia to N-końcowa domena lektyny. Selektyny L, P, E różnią się liczbą bloków w białku regulatorowym dopełniacza. Lektyny to rodzina białek, które specyficznie oddziałują z pewnymi sekwencjami reszt węglowodanowych w składzie glikoprotein, proteoglikanów i glikolipidów macierzy zewnątrzkomórkowej.







 Starszy Makariusz z Egiptu")

- Kościół Wniebowzięcia Najświętszej Marii Panny
- Borys Chramcow. Oczy patrzące w niebo. „Wiedział coś, o czym dawno zapomniałem”
- Grób błogosławionej Aleksandry i ikona Augusta w Darnie
- Świątynia Założenia Szaty na Donskoju
- Świątynia Tichwińskiej Ikony Matki Bożej przy ul.
- Przepis krok po kroku na robienie ciast z ziemniakami w piekarniku
- Placki z piekarnika z ziemniakami
- Przepisy na proste i smaczne sałatki z grzankami
- Przepisy na proste i smaczne sałatki z grzankami
- Zupa rybna Przepisy z okonia morskiego na gotowanie zupy rybnej
- Serca z kurczaka w śmietanie
- Rosnieft – polityka Federalnej Republiki Uralu Kto stanie na czele służby bezpieczeństwa Rosniefti
- Pułkownik Romanow i generał Khorev zostali wyprzedzeni przez dużą redystrybucję celną
- Poważne oskarżenia: kobiety poskarżyły się Matwience i Bastrykinowi na córkę głównego prokuratora wojskowego Fridinskiego Siergieja Nikołajewicza
- Napisz Aleksander Jewgienijewicz Lebiediew
- Castling Shah Galina Dolova przeczytaj w całości
- Recenzja książki: „Kąt widzenia w zeszycie ćwiczeń szybkiego czytania” Tony’ego Buzana
- Wielebny Dawid z Serpuchowa
- Czcigodny Makariusz Wielki, Egipcjanin (†391) Starszy Makariusz z Egiptu
- W czym pomaga ikona „Zdesperowana nadzieja”?